

Errors in the bisulfite conversion of DNA: modulating inappropriate- and failed-conversion frequencies
- PMID: 18984622
- PMCID: PMC2602783
- DOI: 10.1093/nar/gkn691
Errors in the bisulfite conversion of DNA: modulating inappropriate- and failed-conversion frequencies
Erratum in
- Nucleic Acids Res. 2009 Aug;37(15):5235
Abstract
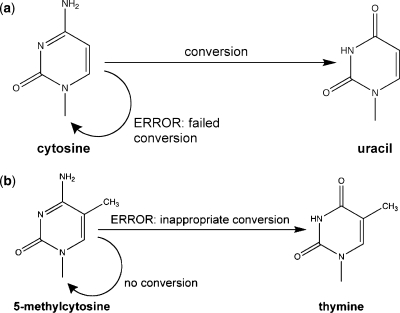
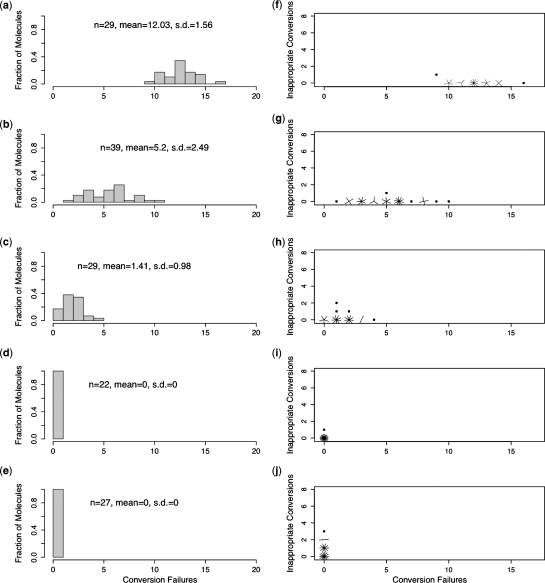
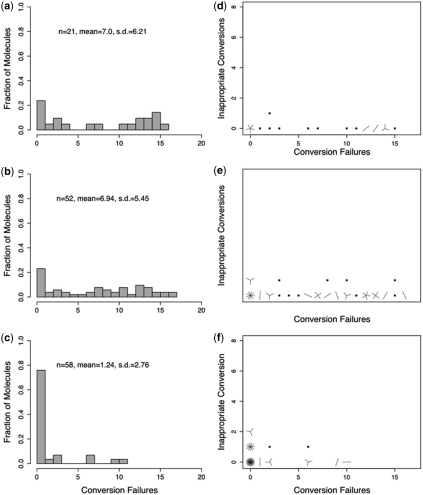
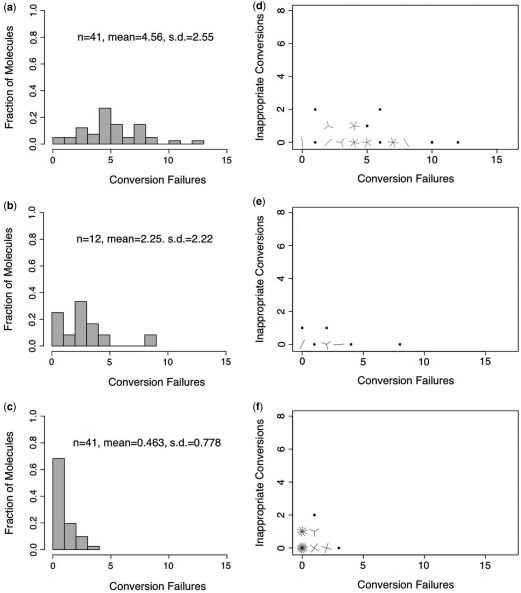
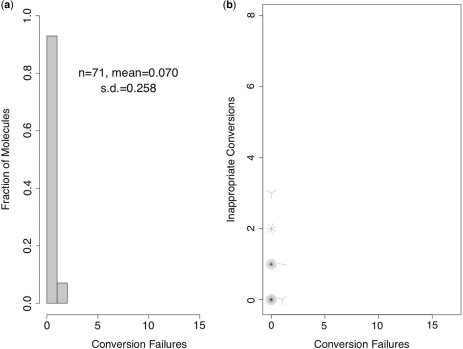
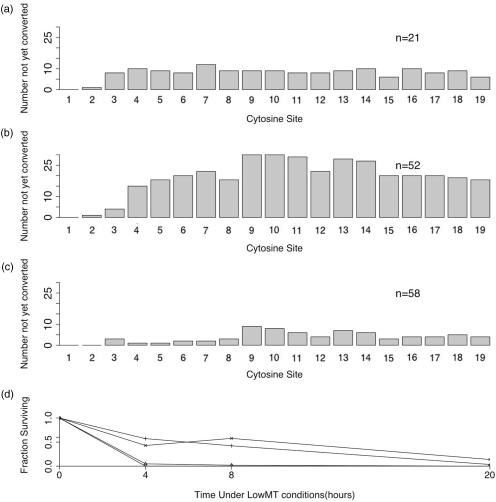
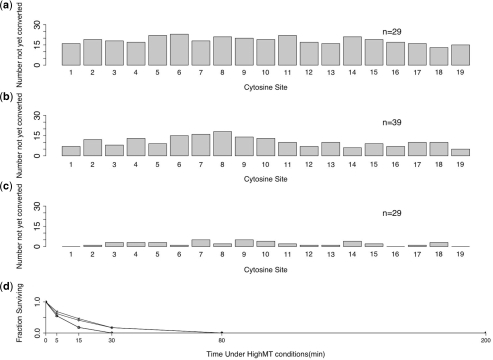
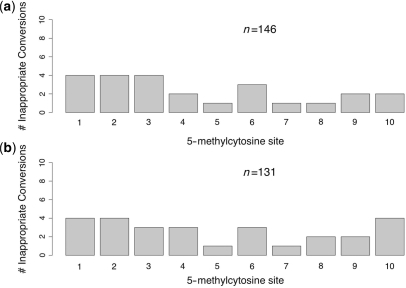
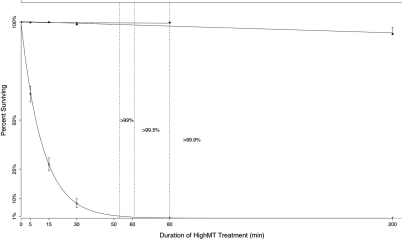
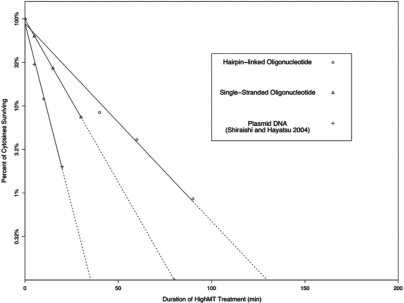
Bisulfite treatment can be used to ascertain the methylation states of individual cytosines in DNA. Ideally, bisulfite treatment deaminates unmethylated cytosines to uracils, and leaves 5-methylcytosines unchanged. Two types of bisulfite-conversion error occur: inappropriate conversion of 5-methylcytosine to thymine, and failure to convert unmethylated cytosine to uracil. Conventional bisulfite treatment requires hours of exposure to low-molarity, low-temperature bisulfite ('LowMT') and, sometimes, thermal denaturation. An alternate, high-molarity, high-temperature ('HighMT') protocol has been reported to accelerate conversion and to reduce inappropriate conversion. We used molecular encoding to obtain validated, individual-molecule data on failed- and inappropriate-conversion frequencies for LowMT and HighMT treatments of both single-stranded and hairpin-linked oligonucleotides. After accounting for bisulfite-independent error, we found that: (i) inappropriate-conversion events accrue predominantly on molecules exposed to bisulfite after they have attained complete or near-complete conversion; (ii) the HighMT treatment is preferable because it yields greater homogeneity among sites and among molecules in conversion rates, and thus yields more reliable data; (iii) different durations of bisulfite treatment will yield data appropriate to address different experimental questions; and (iv) conversion errors can be used to assess the validity of methylation data collected without the benefit of molecular encoding.
Figures
References
-
- Stöger R, Kubicka P, Liu CG, Kafri T, Razin A, Cedar H, Barlow DP. Maternal-specific methylation of the imprinted mouse Igf2r locus identies the expressed locus as carrying the imprinting signal. Cell. 1993;73:61–71. - PubMed
-
- Swain JL, Stewart TA, Leder P. Parental legacy determines methylation and expression of an autosomal transgene: a molecular mechanism for parental imprinting. Cell. 1987;50:719–727. - PubMed
-
- Feinberg AP, Vogelstein B. Alterations in DNA methylation in human colon neoplasia. Semin. Surg. Oncol. 1987;3:149–151. - PubMed
-
- Laird CD, Jaffe E, Karpen G, Lamb M, Nelson R. Fragile sites in human chromosomes as regions of late-replicating DNA. Trends Genet. 1987;3:274–281.
-
- Jacobsen SE, Meyerowitz EM. Hypermethylated SUPERMAN epigenetic alleles in Arabidopsis. Science. 1997;277:1100–1103. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
