

Spatiotemporal regulation of fibroblast growth factor signal blocking for endoderm formation in Xenopus laevis
- PMID: 18985013
- PMCID: PMC2679358
- DOI: 10.3858/emm.2008.40.5.550
Spatiotemporal regulation of fibroblast growth factor signal blocking for endoderm formation in Xenopus laevis
Abstract
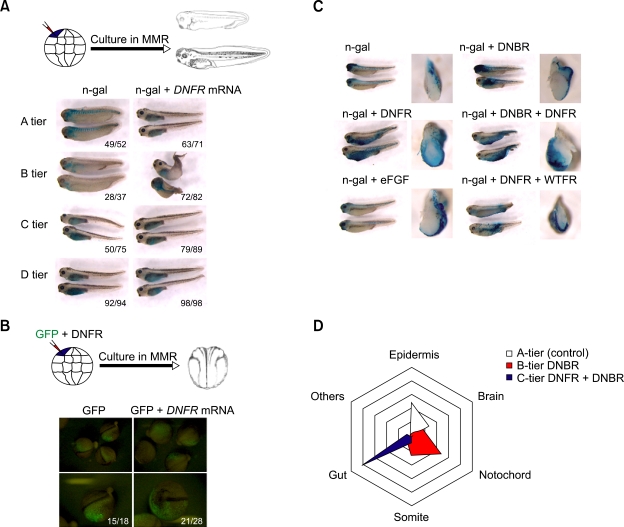
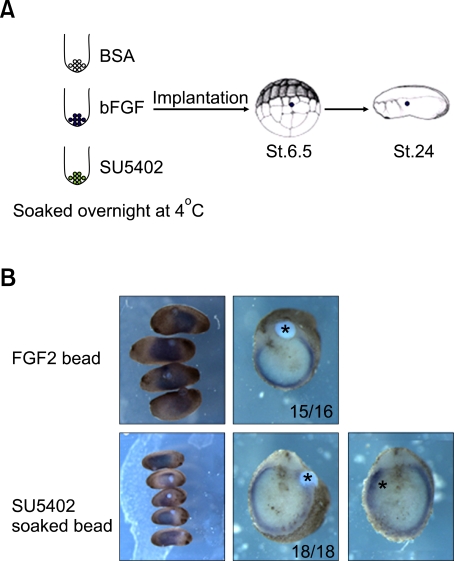
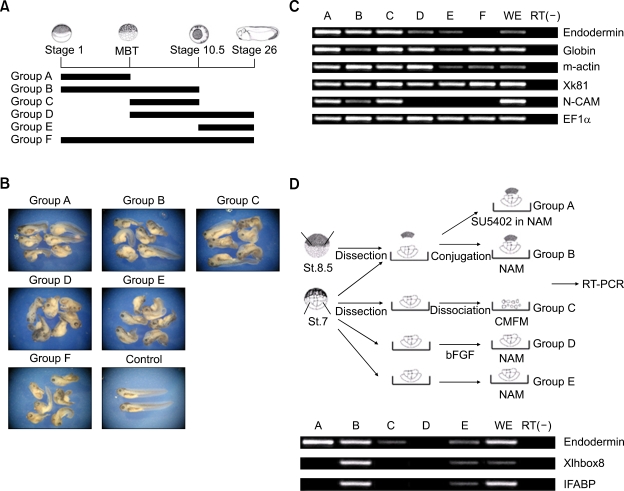
We have previously shown that the inhibition of fibroblast growth factor (FGF) signaling induced endodermal gene expression in the animal cap and caused the expansion of the endodermal mass in Xenopus embryos. However, we still do not know whether or not the alteration of FGF signaling controls embryonic cell fate, or when FGF signal blocking is required for endoderm formation in Xenopus. Here, we show that FGF signal blocking in embryonic cells causes their descendants to move into the endodermal region and to express endodermal genes. It is also interesting that blocking FGF signaling between fertilization and embryonic stage 10.5 promotes endoderm formation, but persistent FGF signaling blocking after stage 10.5 restricts endoderm formation and differentiation.
Figures
References
-
- Amaya E, Stein PA, Musci TJ, Kirschner MW. FGF signalling in the early specification of mesoderm in Xenopus. Development. 1993;118:477–487. - PubMed
-
- Cha SW, Hwang YS, Chae JP, Lee SY, Lee HS, Daar I, Park MJ, Kim J. Inhibition of FGF signaling causes expansion of the endoderm in Xenopus. Biochem Biophys Res Commun. 2004;315:100–106. - PubMed
-
- Ciruna B, Rossant J. FGF signaling regulates mesoderm cell fate specification and morphogenetic movement at the primitive streak. Dev Cell. 2001;1:37–49. - PubMed
-
- Clements D, Friday RV, Woodland HR. Mode of action of VegT in mesoderm and endoderm formation. Development. 1999;126:4903–4911. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous