

Molecular bases for the recognition of short peptide substrates and cysteine-directed modifications of human insulin-degrading enzyme
- PMID: 18986166
- PMCID: PMC2652632
- DOI: 10.1021/bi801192h
Molecular bases for the recognition of short peptide substrates and cysteine-directed modifications of human insulin-degrading enzyme
Abstract
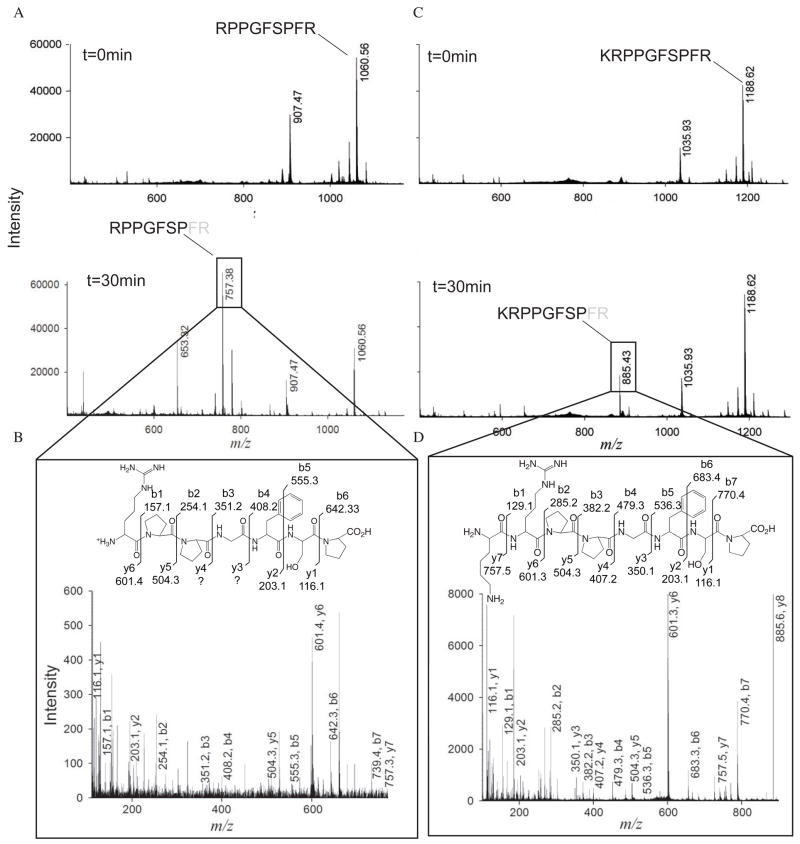
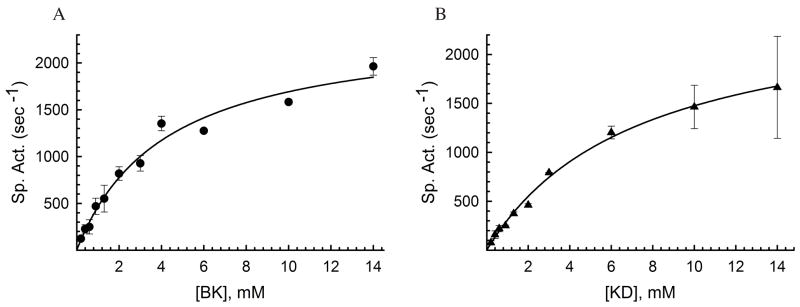
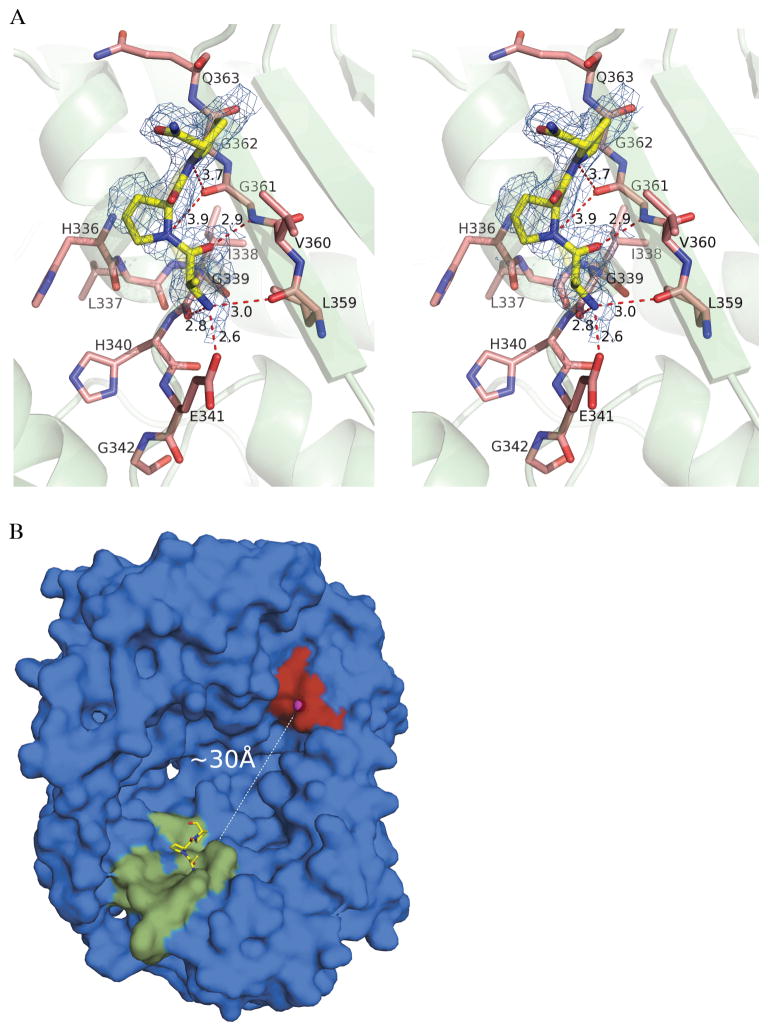
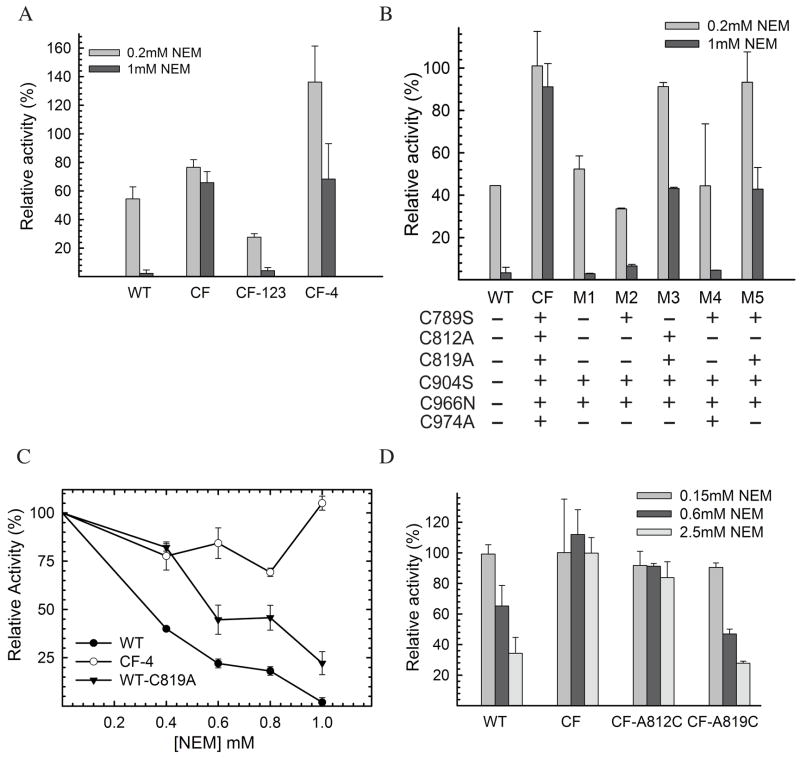
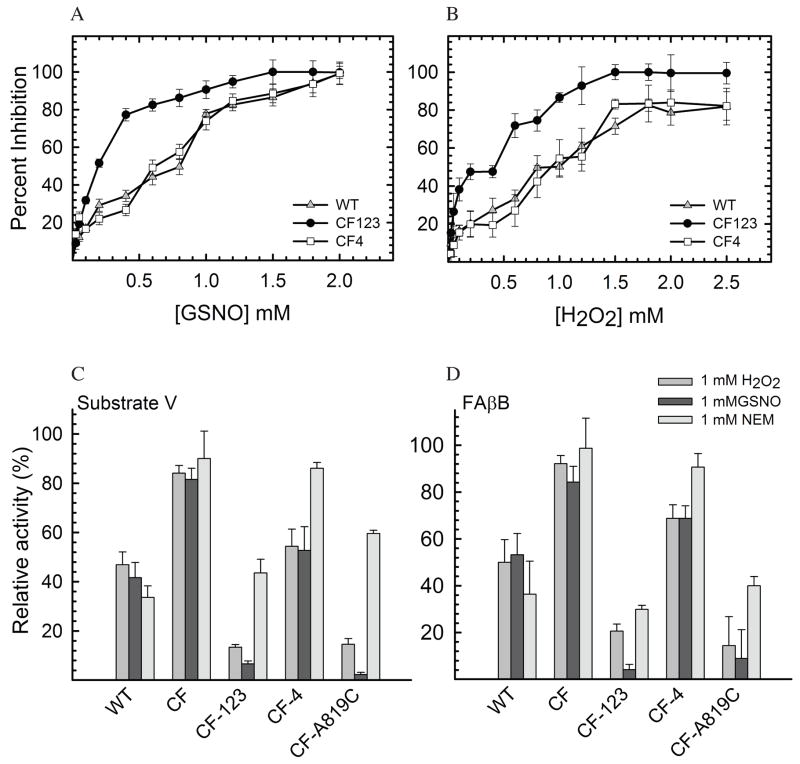
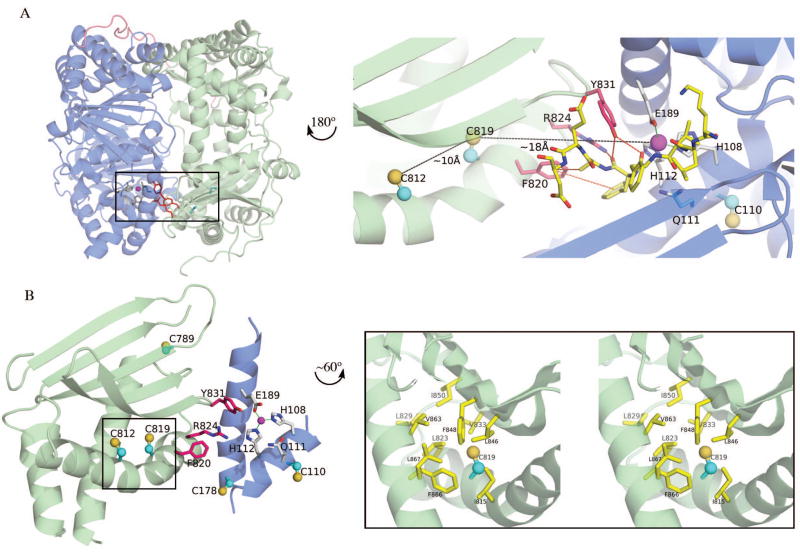
Insulin degrading enzyme (IDE) utilizes a large catalytic chamber to selectively bind and degrade peptide substrates such as insulin and amyloid beta (Abeta). Tight interactions with substrates occur at an exosite located approximately 30 A away from the catalytic center that anchors the N-terminus of substrates to facilitate binding and subsequent cleavages at the catalytic site. However, IDE also degrades peptide substrates that are too short to occupy both the catalytic site and the exosite simultaneously. Here, we use kinins as a model system to address the kinetics and regulation of human IDE with short peptides. IDE specifically degrades bradykinin and kallidin at the Pro/Phe site. A 1.9 A crystal structure of bradykinin-bound IDE reveals the binding of bradykinin to the exosite and not to the catalytic site. In agreement with observed high K(m) values, this suggests low affinity of bradykinin for IDE. This structure also provides the molecular basis on how the binding of short peptides at the exosite could regulate substrate recognition. We also found that human IDE is potently inhibited by physiologically relevant concentrations of S-nitrosylation and oxidation agents. Cysteine-directed modifications play a key role, since an IDE mutant devoid of all 13 cysteines is insensitive to the inhibition by S-nitrosoglutathione, hydrogen peroxide, or N-ethylmaleimide. Specifically, cysteine 819 of human IDE is located inside the catalytic chamber pointing toward an extended hydrophobic pocket and is critical for the inactivation. Thiol-directed modification of this residue likely causes local structural perturbation to reduce substrate binding and catalysis.
Figures
Similar articles
-
Molecular basis for the recognition and cleavages of IGF-II, TGF-alpha, and amylin by human insulin-degrading enzyme.J Mol Biol. 2010 Jan 15;395(2):430-43. doi: 10.1016/j.jmb.2009.10.072. Epub 2009 Nov 5. J Mol Biol. 2010. PMID: 19896952 Free PMC article.
-
Structure based discovery of small molecules to regulate the activity of human insulin degrading enzyme.PLoS One. 2012;7(2):e31787. doi: 10.1371/journal.pone.0031787. Epub 2012 Feb 15. PLoS One. 2012. PMID: 22355395 Free PMC article.
-
Molecular basis of catalytic chamber-assisted unfolding and cleavage of human insulin by human insulin-degrading enzyme.J Biol Chem. 2009 May 22;284(21):14177-88. doi: 10.1074/jbc.M900068200. Epub 2009 Mar 25. J Biol Chem. 2009. PMID: 19321446 Free PMC article.
-
Insulin-degrading enzyme: structure-function relationship and its possible roles in health and disease.Curr Pharm Des. 2009;15(31):3644-55. doi: 10.2174/138161209789271799. Curr Pharm Des. 2009. PMID: 19925417 Review.
-
Amyloid beta-degrading cryptidases: insulin degrading enzyme, presequence peptidase, and neprilysin.Cell Mol Life Sci. 2008 Aug;65(16):2574-85. doi: 10.1007/s00018-008-8112-4. Cell Mol Life Sci. 2008. PMID: 18470479 Free PMC article. Review.
Cited by
-
Aberrant protein s-nitrosylation in neurodegenerative diseases.Neuron. 2013 May 22;78(4):596-614. doi: 10.1016/j.neuron.2013.05.005. Neuron. 2013. PMID: 23719160 Free PMC article. Review.
-
Cysteine 904 is required for maximal insulin degrading enzyme activity and polyanion activation.PLoS One. 2012;7(10):e46790. doi: 10.1371/journal.pone.0046790. Epub 2012 Oct 15. PLoS One. 2012. PMID: 23077523 Free PMC article.
-
Insulin-degrading enzyme prevents α-synuclein fibril formation in a nonproteolytical manner.Sci Rep. 2015 Jul 31;5:12531. doi: 10.1038/srep12531. Sci Rep. 2015. PMID: 26228656 Free PMC article.
-
In vitro degradation of insulin-like peptide 3 by insulin-degrading enzyme.Protein J. 2010 Feb;29(2):93-8. doi: 10.1007/s10930-009-9226-8. Protein J. 2010. PMID: 20082125
-
Designed inhibitors of insulin-degrading enzyme regulate the catabolism and activity of insulin.PLoS One. 2010 May 7;5(5):e10504. doi: 10.1371/journal.pone.0010504. PLoS One. 2010. PMID: 20498699 Free PMC article.
References
-
- Duckworth WC, Bennett RG, Hamel FG. Insulin degradation: progress and potential. Endocr Rev. 1998;19:608–624. - PubMed
-
- Kurochkin IV. Insulin-degrading enzyme: embarking on amyloid destruction. Trends Biochem Sci. 2001;26:421–425. - PubMed
-
- Mirsky IA, Broth-Kahn RH. The inactivation of insulin by tissue extracts. I. The distribution and properties of insulin inactivating extracts (insulinase) Arch Biochem. 1949;20:1–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
