

Evolution acts on enhancer organization to fine-tune gradient threshold readouts
- PMID: 18986212
- PMCID: PMC2577699
- DOI: 10.1371/journal.pbio.0060263
Evolution acts on enhancer organization to fine-tune gradient threshold readouts
Abstract
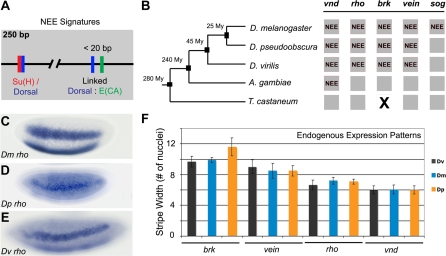
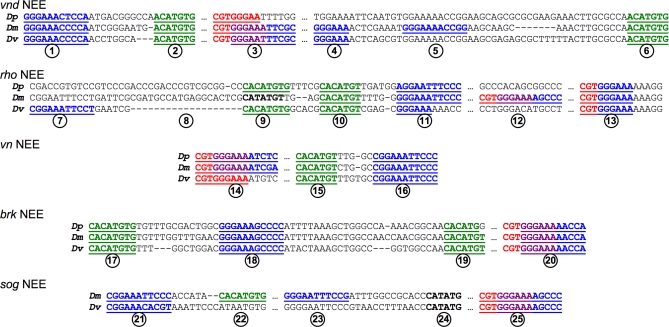
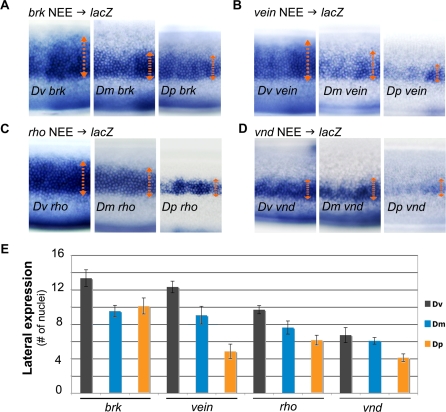
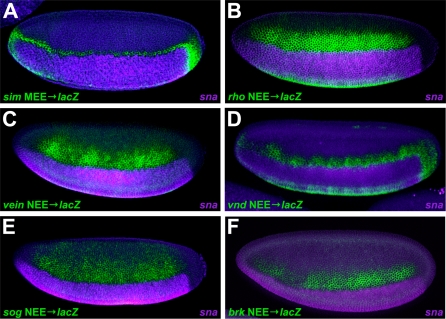
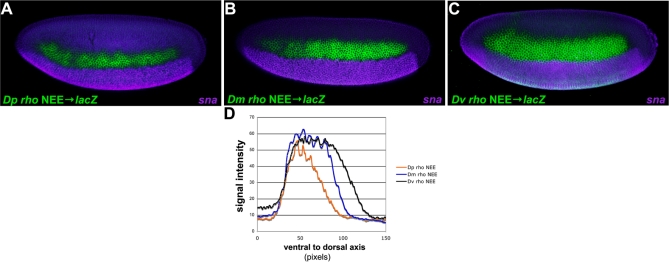
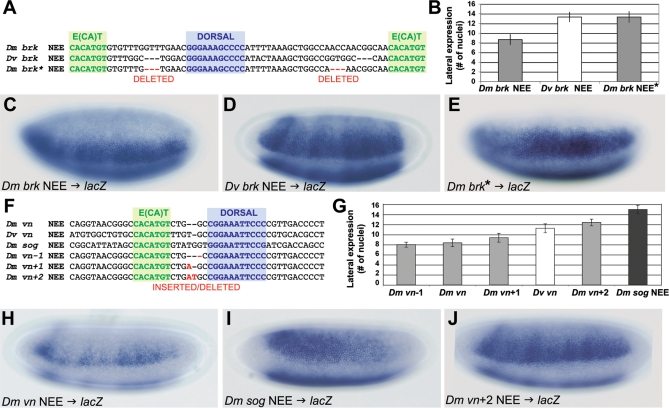
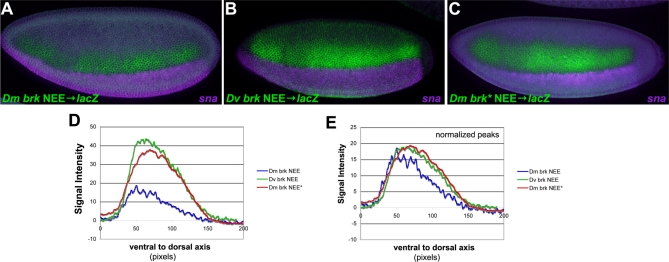
The elucidation of principles governing evolution of gene regulatory sequence is critical to the study of metazoan diversification. We are therefore exploring the structure and organizational constraints of regulatory sequences by studying functionally equivalent cis-regulatory modules (CRMs) that have been evolving in parallel across several loci. Such an independent dataset allows a multi-locus study that is not hampered by nonfunctional or constrained homology. The neurogenic ectoderm enhancers (NEEs) of Drosophila melanogaster are one such class of coordinately regulated CRMs. The NEEs share a common organization of binding sites and as a set would be useful to study the relationship between CRM organization and CRM activity across evolving lineages. We used the D. melanogaster transgenic system to screen for functional adaptations in the NEEs from divergent drosophilid species. We show that the individual NEE modules across a genome in any one lineage have independently evolved adaptations to compensate for lineage-specific developmental and/or genomic changes. Specifically, we show that both the site composition and the site organization of NEEs have been finely tuned by distinct, lineage-specific selection pressures in each of the three divergent species that we have examined: D. melanogaster, D. pseudoobscura, and D. virilis. Furthermore, by precisely altering the organization of NEEs with different morphogen gradient threshold readouts, we show that CRM organizational evolution is sufficient for explaining changes in enhancer activity. Thus, evolution can act on CRM organization to fine-tune morphogen gradient threshold readouts over a wide dynamic range. Our study demonstrates that equivalence classes of CRMs are powerful tools for detecting lineage-specific adaptations by gene regulatory sequences.
Conflict of interest statement
Figures
Similar articles
-
Functional analysis of eve stripe 2 enhancer evolution in Drosophila: rules governing conservation and change.Development. 1998 Mar;125(5):949-58. doi: 10.1242/dev.125.5.949. Development. 1998. PMID: 9449677
-
Molecular dissection of cis-regulatory modules at the Drosophila bithorax complex reveals critical transcription factor signature motifs.Dev Biol. 2011 Nov 15;359(2):290-302. doi: 10.1016/j.ydbio.2011.07.028. Epub 2011 Jul 28. Dev Biol. 2011. PMID: 21821017 Free PMC article.
-
Gain of cis-regulatory activities underlies novel domains of wingless gene expression in Drosophila.Proc Natl Acad Sci U S A. 2015 Jun 16;112(24):7524-9. doi: 10.1073/pnas.1509022112. Epub 2015 Jun 1. Proc Natl Acad Sci U S A. 2015. PMID: 26034272 Free PMC article.
-
Tracking adaptive evolutionary events in genomic sequences.Genome Biol. 2002;3(6):REVIEWS1018. doi: 10.1186/gb-2002-3-6-reviews1018. Epub 2002 May 29. Genome Biol. 2002. PMID: 12093382 Free PMC article. Review.
-
Dissecting the regulatory switches of development: lessons from enhancer evolution in Drosophila.Development. 2010 Jan;137(1):5-13. doi: 10.1242/dev.036160. Development. 2010. PMID: 20023155 Free PMC article. Review.
Cited by
-
Deciphering a transcriptional regulatory code: modeling short-range repression in the Drosophila embryo.Mol Syst Biol. 2010;6:341. doi: 10.1038/msb.2009.97. Epub 2010 Jan 19. Mol Syst Biol. 2010. PMID: 20087339 Free PMC article.
-
Distinct functional constraints partition sequence conservation in a cis-regulatory element.PLoS Genet. 2011 Jun;7(6):e1002095. doi: 10.1371/journal.pgen.1002095. Epub 2011 Jun 2. PLoS Genet. 2011. PMID: 21655084 Free PMC article.
-
Mechanisms of scaling in pattern formation.Development. 2013 Dec;140(24):4830-43. doi: 10.1242/dev.100511. Development. 2013. PMID: 24301464 Free PMC article. Review.
-
Low affinity binding site clusters confer hox specificity and regulatory robustness.Cell. 2015 Jan 15;160(1-2):191-203. doi: 10.1016/j.cell.2014.11.041. Epub 2014 Dec 31. Cell. 2015. PMID: 25557079 Free PMC article.
-
Dissecting sources of quantitative gene expression pattern divergence between Drosophila species.Mol Syst Biol. 2012;8:604. doi: 10.1038/msb.2012.35. Mol Syst Biol. 2012. PMID: 22893002 Free PMC article.
References
-
- Arnone MI, Davidson EH. The hardwiring of development: organization and function of genomic regulatory systems. Development. 1997;124:1851–1864. - PubMed
-
- Gluzman Y, Shenk T. Enhancers and eukaryotic gene expression. Cold Spring Harbor: Cold Spring Harbor Laboratory; 1983. 218
-
- Small S, Kraut R, Hoey T, Warrior R, Levine M. Transcriptional regulation of a pair-rule stripe in Drosophila. Genes Dev. 1991;5:827–839. - PubMed
-
- Uchikawa M, Ishida Y, Takemoto T, Kamachi Y, Kondoh H. Functional analysis of chicken Sox2 enhancers highlights an array of diverse regulatory elements that are conserved in mammals. Dev Cell. 2003;4:509–519. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
