

Hey1 basic helix-loop-helix protein plays an important role in mediating BMP9-induced osteogenic differentiation of mesenchymal progenitor cells
- PMID: 18986983
- PMCID: PMC2610517
- DOI: 10.1074/jbc.M806389200
Hey1 basic helix-loop-helix protein plays an important role in mediating BMP9-induced osteogenic differentiation of mesenchymal progenitor cells
Abstract
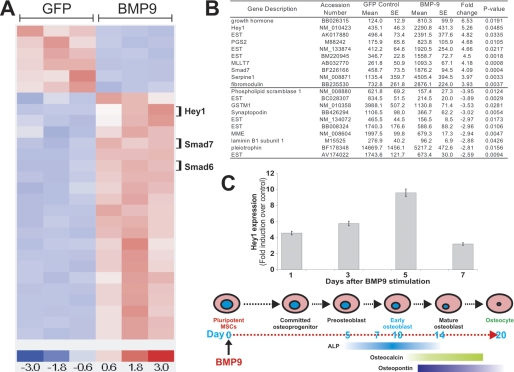
Pluripotent mesenchymal stem cells (MSCs) are bone marrow stromal progenitor cells that can differentiate into osteogenic, chondrogenic, adipogenic, and myogenic lineages. We previously demonstrated that bone morphogenetic protein (BMP) 9 is one of the most potent and yet least characterized BMPs that are able to induce osteogenic differentiation of MSCs both in vitro and in vivo. Here, we conducted gene expression-profiling analysis and identified that Hey1 of the hairy/Enhancer of split-related repressor protein basic helix-loop-helix family was among the most significantly up-regulated early targets in BMP9-stimulated MSCs. We demonstrated that Hey1 expression was up-regulated at the immediate early stage of BMP9-induced osteogenic differentiation. Chromatin immunoprecipitation analysis indicated that Hey1 may be a direct target of the BMP9-induced Smad signaling pathway. Silencing Hey1 expression diminished BMP9-induced osteogenic differentiation both in vitro and in vivo and led to chondrogenic differentiation. Likewise, constitutive Hey1 expression augmented BMP9-mediated bone matrix mineralization. Hey1 and Runx2 were shown to act synergistically in BMP9-induced osteogenic differentiation, and Runx2 expression significantly decreased in the absence of Hey1, suggesting that Runx2 may function downstream of Hey1. Accordingly, the defective osteogenic differentiation caused by Hey1 knockdown was rescued by exogenous Runx2 expression. Thus, our findings suggest that Hey1, through its interplay with Runx2, may play an important role in regulating BMP9-induced osteoblast lineage differentiation of MSCs.
Figures





References
-
- Pittenger, M. F., Mackay, A. M., Beck, S. C., Jaiswal, R. K., Douglas, R., Mosca, J. D., Moorman, M. A., Simonetti, D. W., Craig, S., and Marshak, D. R. (1999) Science 284 143-147 - PubMed
-
- Prockop, D. J. (1997) Science 276 71-74 - PubMed
-
- Aubin, J. E. (1998) J. Cell. Biochem. supplemental 31 73-82 - PubMed
-
- Ducy, P., Schinke, T., and Karsenty, G. (2000) Science 289 1501-1504 - PubMed
-
- Caplan, A. I., and Bruder, S. P. (2001) Trends Mol. Med. 7 259-264 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
