

The genesis of cartilage size and shape during development and evolution
- PMID: 18987028
- PMCID: PMC2836324
- DOI: 10.1242/dev.023309
The genesis of cartilage size and shape during development and evolution
Abstract
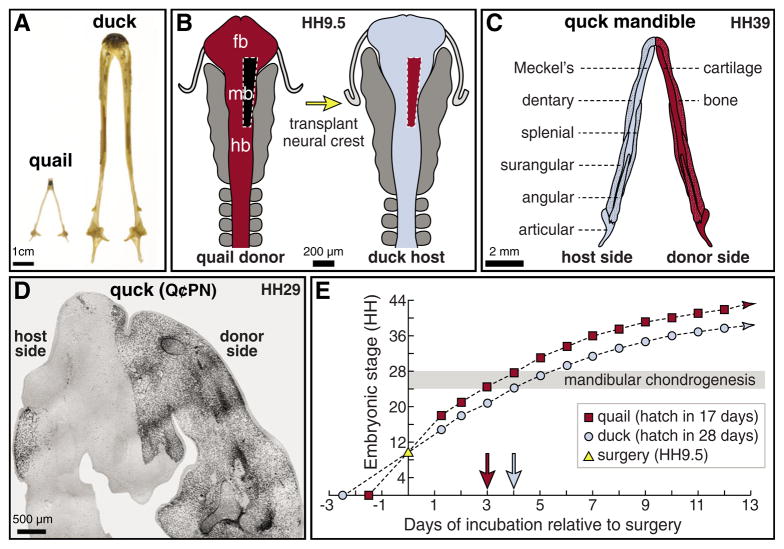
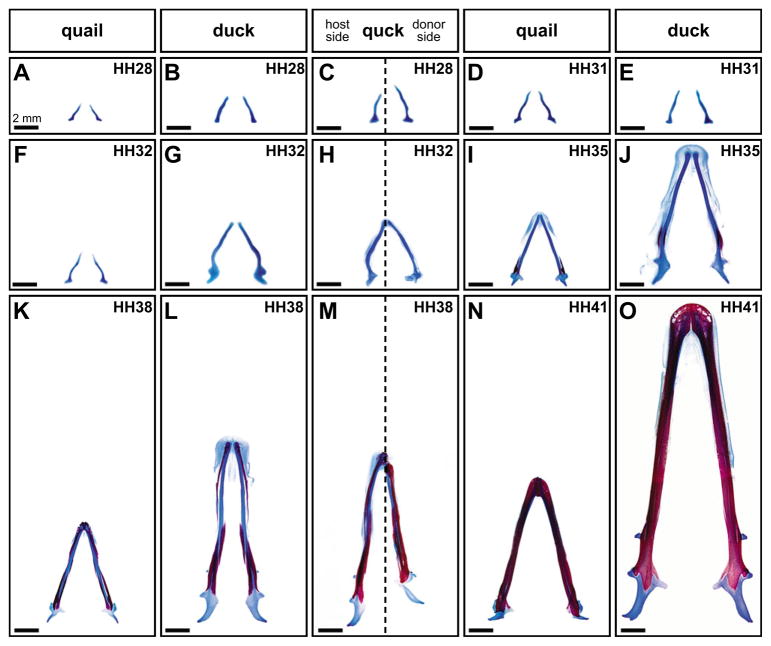
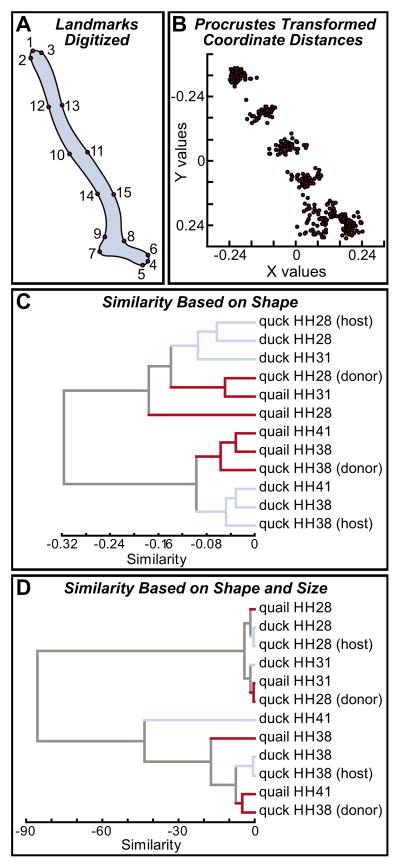
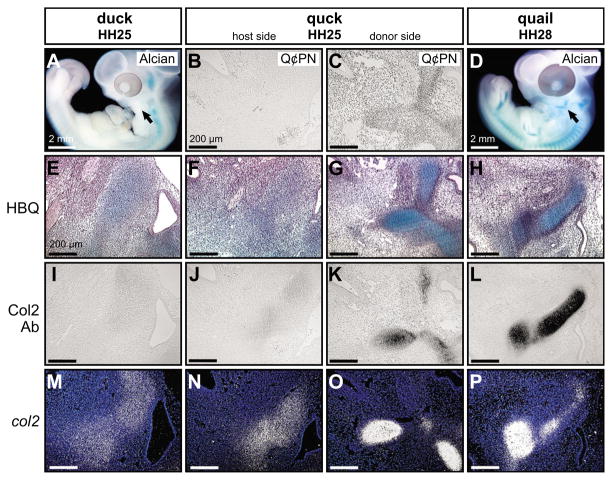
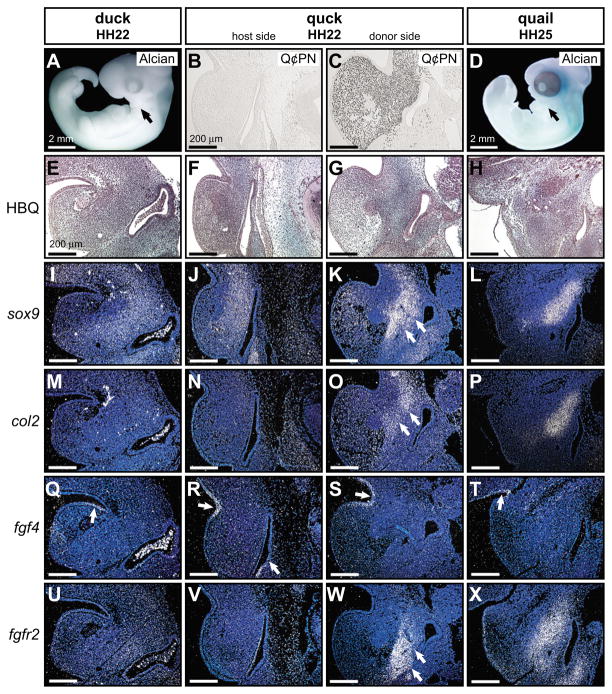
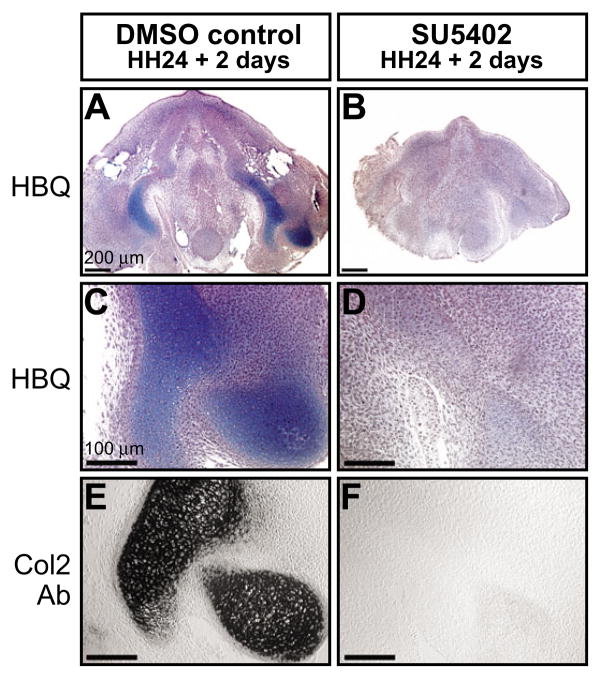
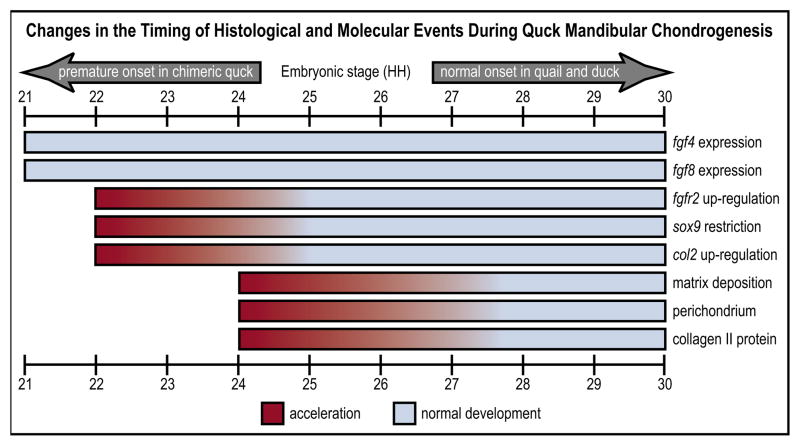
How do cartilaginous elements attain their characteristic size and shape? Two intimately coupled processes underlie the patterned growth of cartilage. The first is histogenesis, which entails the production of cartilage as a discrete tissue; the second is morphogenesis, which pertains to the origins of three-dimensional form. Histogenesis relies on cues that promote the chondrogenic differentiation of mesenchymal cells, whereas morphogenesis requires information that imbues cartilage with stage-specific (e.g. embryonic versus adult), region-specific (e.g. cranial versus appendicular) and species-specific size and shape. Previous experiments indicate that early programmatic events and subsequent signaling interactions enable chondrogenic mesenchyme to undergo histogenesis and morphogenesis, but precise molecular and cellular mechanisms that generate cartilage size and shape remain unclear. In the face and jaws, neural crest-derived mesenchyme clearly plays an important role, given that this embryonic population serves as the source of chondrocytes and of species-specific patterning information. To elucidate mechanisms through which neural crest-derived mesenchyme affects cartilage size and shape, we made chimeras using quail and duck embryos, which differ markedly in their craniofacial anatomy and rates of maturation. Transplanting neural crest cells from quail to duck demonstrates that mesenchyme imparts both stage-specific and species-specific size and shape to cartilage by controlling the timing of preceding and requisite molecular and histogenic events. In particular, we find that mesenchyme regulates FGF signaling and the expression of downstream effectors such as sox9 and col2a1. The capacity of neural crest-derived mesenchyme to orchestrate spatiotemporal programs for chondrogenesis autonomously, and to implement cartilage size and shape across embryonic stages and between species simultaneously, provides a novel mechanism linking ontogeny and phylogeny.
Figures
References
-
- Abzhanov A, Tabin CJ. Shh and Fgf8 act synergistically to drive cartilage outgrowth during cranial development. Dev Biol. 2004;273:134–148. - PubMed
-
- Abzhanov A, Protas M, Grant BR, Grant PR, Tabin CJ. Bmp4 and morphological variation of beaks in Darwin’s finches. Science. 2004;305:1462–1465. - PubMed
-
- Alberch P, Gould SJ, Oster GF, Wake DB. Size and shape in ontogeny and phylogeny. Paleobiology. 1979;5:296–317.
-
- Albrecht UEG, Helms JA, Lin H. Visualization of gene expression patterns by in situ hybridization. In: Daston GP, editor. Molecular and Cellular Methods in Developmental Toxicology. Boca Raton, FL: CRC Press; 1997. pp. 23–48.
-
- Andres G. Untersuchungen an Chimären von Triton und Bombinator. Genetica. 1949;24:387–534.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
