

Characterization of large conformational changes and autoproteolysis in the maturation of a T=4 virus capsid
- PMID: 18987141
- PMCID: PMC2612371
- DOI: 10.1128/JVI.01859-08
Characterization of large conformational changes and autoproteolysis in the maturation of a T=4 virus capsid
Abstract
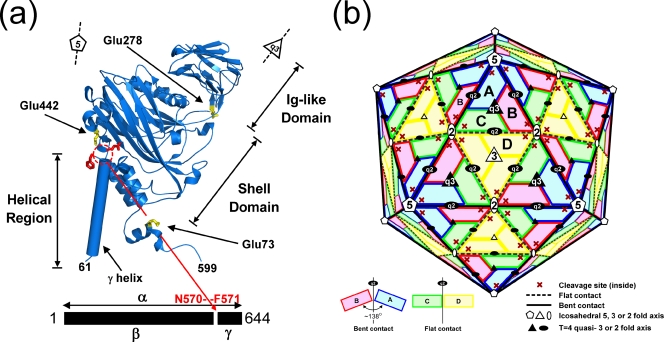
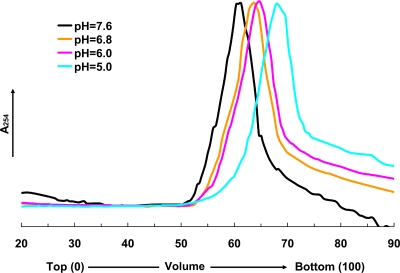
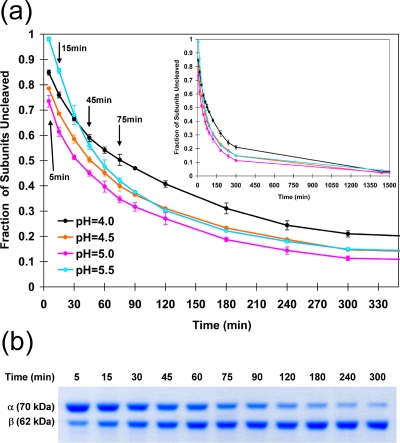
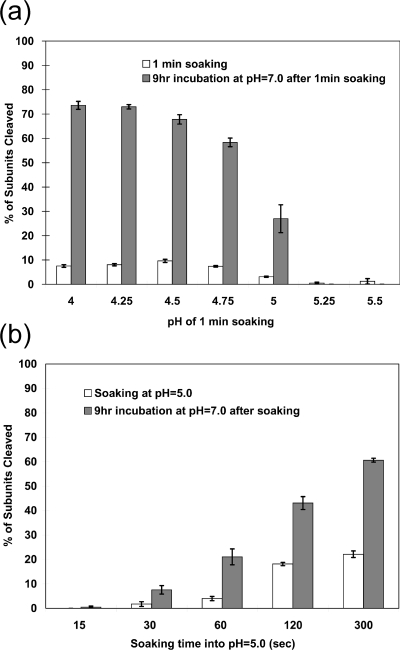
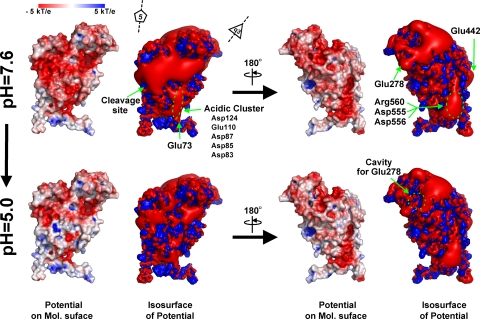
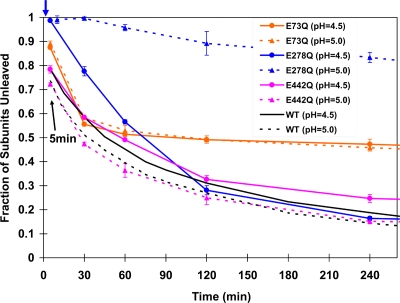
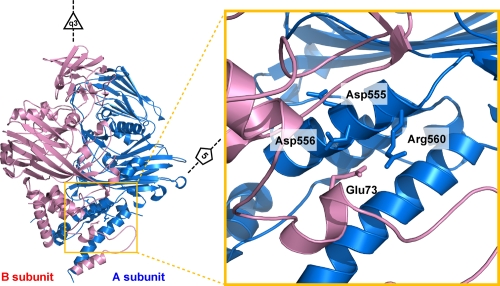
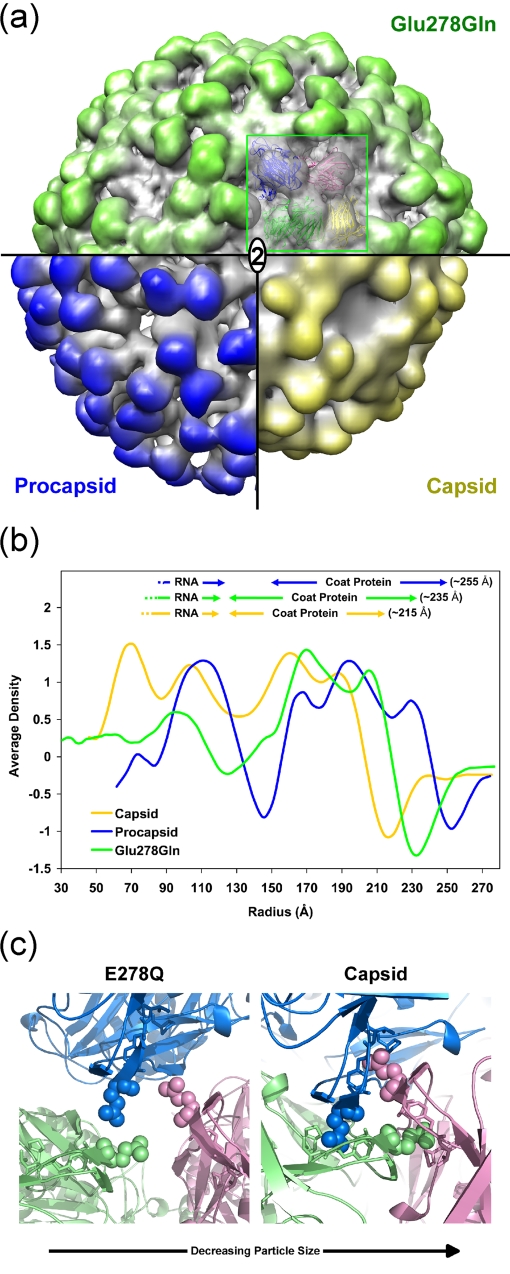
Nudaurelia capensis omega virus-like particles have been characterized as a 480-A procapsid and a 410-A capsid, both with T=4 quasisymmetry. Procapsids transition to capsids when pH is lowered from 7.6 to 5.0. Capsids undergo autoproteolysis at residue 570, generating the 74-residue C-terminal polypeptide that remains with the particle. Here we show that the particle size becomes smaller under conditions between pH 6.8 and 6.0 without activating cleavage and that the particle remains at an intermediate size when the pH is carefully maintained. At pH 5.8, cleavage is very slow, becoming detectable only after 9 h. The optimum pH for cleavage is 5.0 (half-life, approximately 30 min), with a significant reduction in the cleavage rate at pH values below 5. We also show that lowering the pH is required only to make the virus particles compact and to presumably form the active site for autoproteolysis but not for the chemistry of cleavage. The cleavage reaction proceeds at pH 7.0 after approximately 10% of the subunits cleave at pH 5.0. Employing the virion crystal structure for reference, we investigated the role of electrostatic repulsion of acidic residues in the pH-dependent large conformational changes. Three mutations of Glu to Gln that formed procapsids showed three different phenotypes on maturation. One, close to the threefold and quasithreefold symmetry axes and far from the cleavage site, did not mature at pH 5, and electron cryomicroscopy reconstruction showed that it was intermediate in size between those of the procapsid and capsid; one near the cleavage site exhibited a wild-type phenotype; and a third, far from the cleavage site, resulted in cleavage of 50% of the subunits after 4 h, suggesting quasiequivalent specificity of the mutation.
Figures
References
-
- Agrawal, D. K., and J. E. Johnson. 1995. Assembly of the T = 4 Nudaurelia capensis omega virus capsid protein, post-translational cleavage, and specific encapsidation of its mRNA in a baculovirus expression system. Virology 20789-97. - PubMed
-
- Agrawal, D. K., and J. E. Johnson. 1992. Sequence and analysis of the capsid protein of Nudaurelia capensis omega virus, an insect virus with T = 4 icosahedral symmetry. Virology 190806-814. - PubMed
-
- Brooks, E. M., K. H. Gordon, S. J. Dorrian, E. R. Hines, and T. N. Hanzlik. 2002. Infection of its lepidopteran host by the Helicoverpa armigera stunt virus (Tetraviridae). J. Invertebr. Pathol. 8097-111. - PubMed
-
- Brünger, A. T., P. D. Adams, G. M. Clore, W. L. DeLano, P. Gros, R. W. Grosse-Kunstleve, J. S. Jiang, J. Kuszewski, M. Nilges, N. S. Pannu, R. J. Read, L. M. Rice, T. Simonson, and G. L. Warren. 1998. Crystallography & NMR system: a new software suite for macromolecular structure determination. Acta Crystallogr. D Biol. Crystallogr. 54905-921. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
