

Identification of the LWYIK motif located in the human immunodeficiency virus type 1 transmembrane gp41 protein as a distinct determinant for viral infection
- PMID: 18987155
- PMCID: PMC2612407
- DOI: 10.1128/JVI.01088-08
Identification of the LWYIK motif located in the human immunodeficiency virus type 1 transmembrane gp41 protein as a distinct determinant for viral infection
Abstract
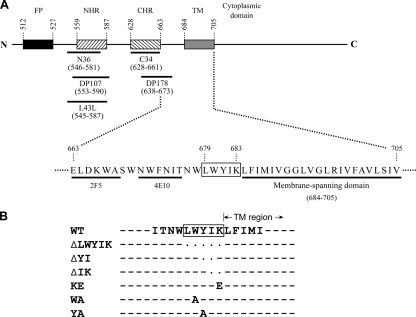
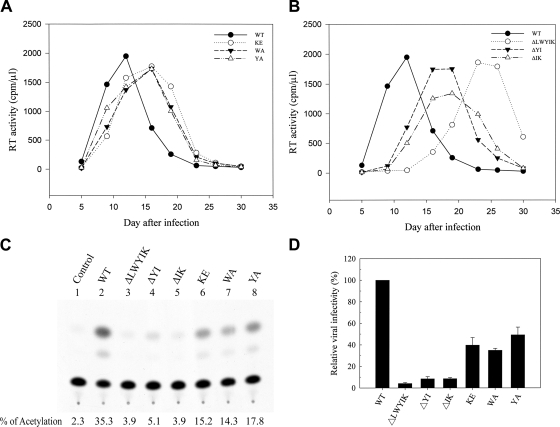
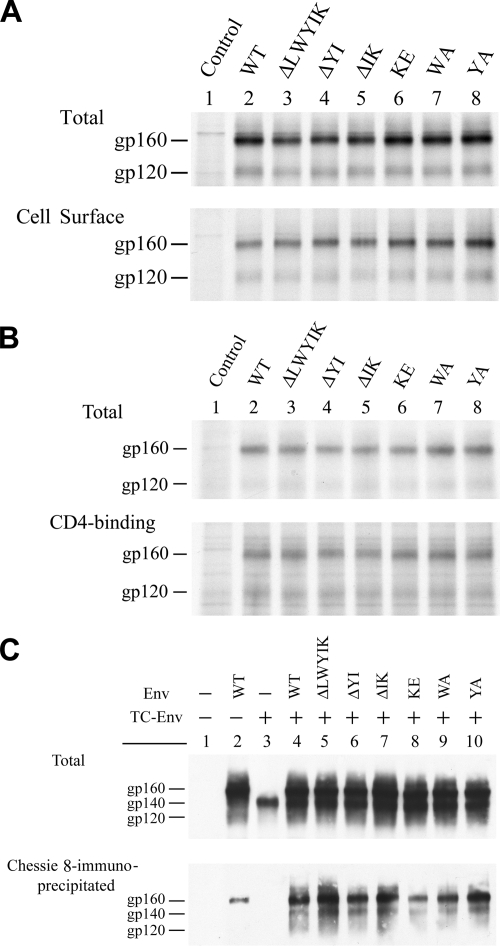
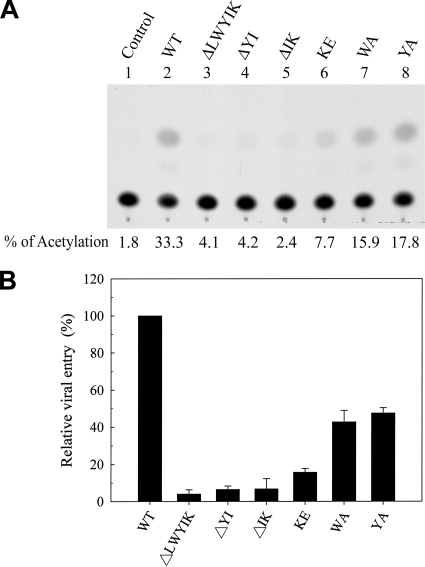
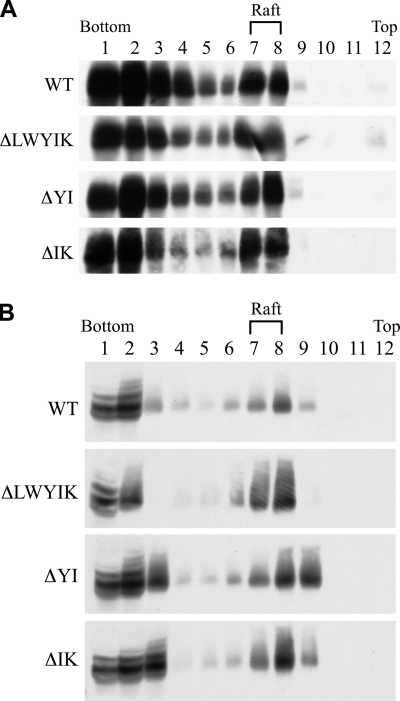
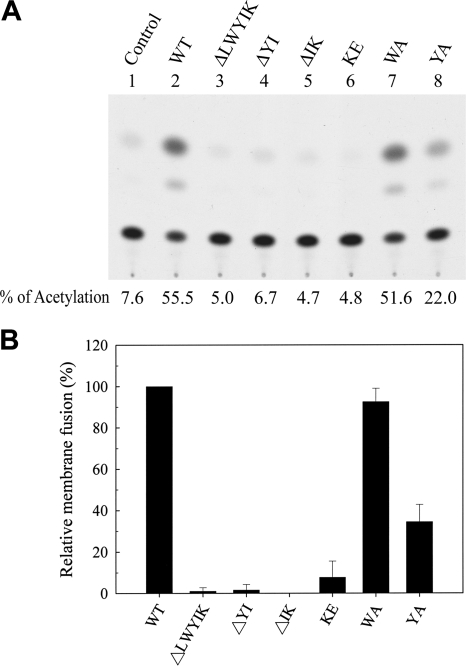
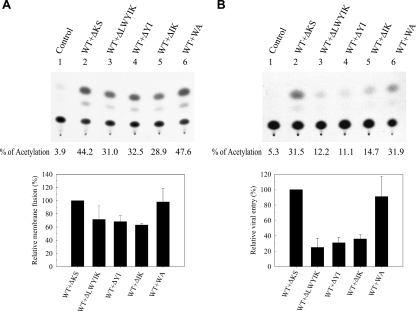
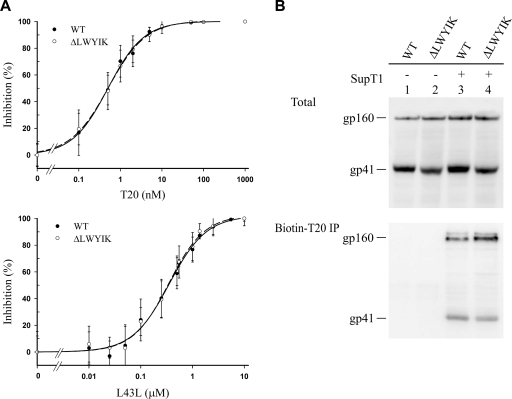
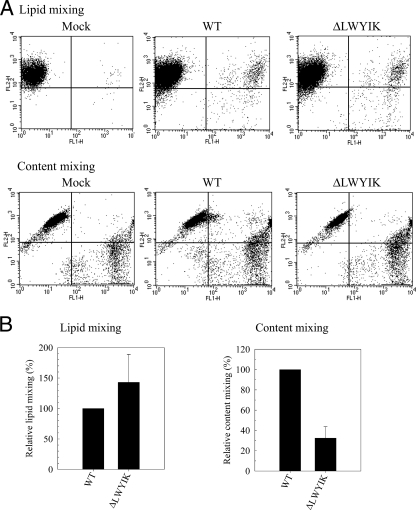
The highly conserved LWYIK motif located immediately proximal to the membrane-spanning domain of the gp41 transmembrane protein of human immunodeficiency virus type 1 has been proposed as being important for the surface envelope (Env) glycoprotein's association with lipid rafts and gp41-mediated membrane fusion. Here we employed substitution and deletion mutagenesis to understand the role of this motif in the virus life cycle. None of the mutants examined affected the synthesis, precursor processing, CD4 binding, oligomerization, or cell surface expression of the Env, nor did they alter Env incorporation into the virus. All of the mutants, particularly the DeltaYI, DeltaIK, and DeltaLWYIK mutants, in which the indicated residues were deleted, exhibited greatly reduced one-cycle viral replication and the Env trans-complementation ability. All of these deletion mutant proteins were still localized in the lipid rafts. With the exception of the Trp-to-Ala (WA) mutant, which exhibited reduced viral infectivity albeit with normal membrane fusion, all mutants displayed loss of some or almost all of the membrane fusion ability. Although these deletion mutants partially inhibited in trans wild-type (WT) Env-mediated fusion, they were more effective in dominantly interfering with WT Env-mediated viral entry when coexpressed with the WT Env, implying a role of this motif in postfusion events as well. Both T20 and L43L peptides derived from the two gp41 extracellular C- and N-terminal alpha-helical heptad repeats, respectively, inhibited WT and DeltaLWYIK Env-mediated viral entry with comparable efficacies. Biotin-tagged T20 effectively captured both the fusion-active, prehairpin intermediates of WT and mutant gp41 upon CD4 activation. Env without the deletion of the LWYIK motif still effectively mediated lipid mixing but inhibited content mixing. Our study demonstrates that the immediate membrane-proximal LWYIK motif acts as a unique and distinct determinant located in the gp41 C-terminal ectodomain by promoting enlargement of fusion pores and postfusion activities.
Figures
References
-
- Anderson, R. G. 1998. The caveolae membrane system. Annu. Rev. Biochem. 67199-225. - PubMed
-
- Bellamy-McIntyre, A. K., C. S. Lay, S. Baar, A. L. Maerz, G. H. Talbo, H. E. Drummer, and P. Poumbourios. 2007. Functional links between the fusion peptide-proximal polar segment and membrane-proximal region of human immunodeficiency virus gp41 in distinct phases of membrane fusion. J. Biol. Chem. 28223104-23116. - PubMed
-
- Brown, D. A., and J. K. Rose. 1992. Sorting of GPI-anchored proteins to glycolipid-enriched membrane subdomains during transport to the apical cell surface. Cell 68533-544. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
