

A unique cell division machinery in the Archaea
- PMID: 18987308
- PMCID: PMC2596248
- DOI: 10.1073/pnas.0809467105
A unique cell division machinery in the Archaea
Abstract
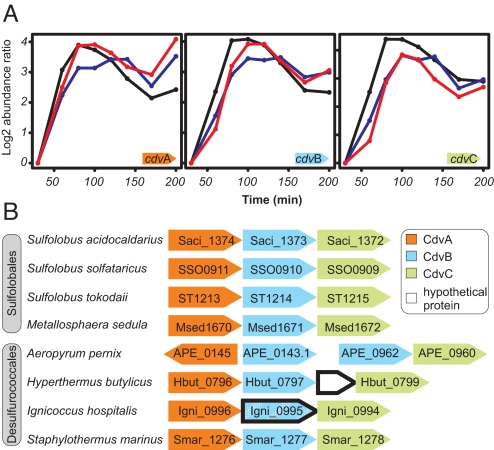
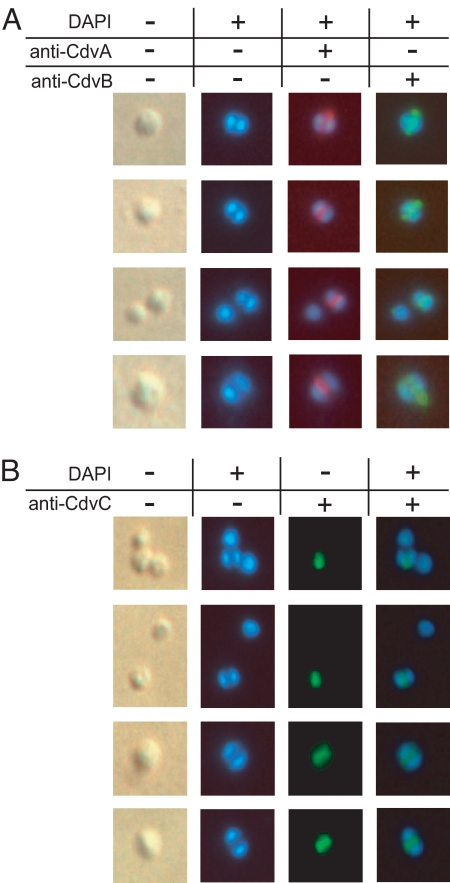
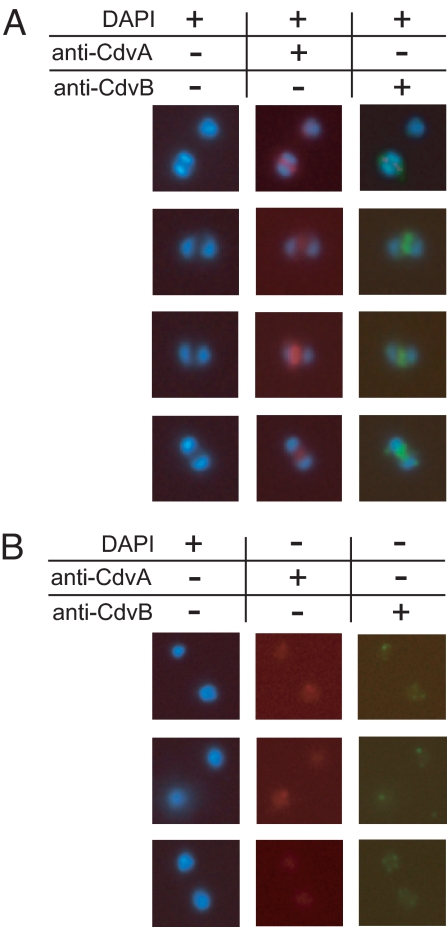
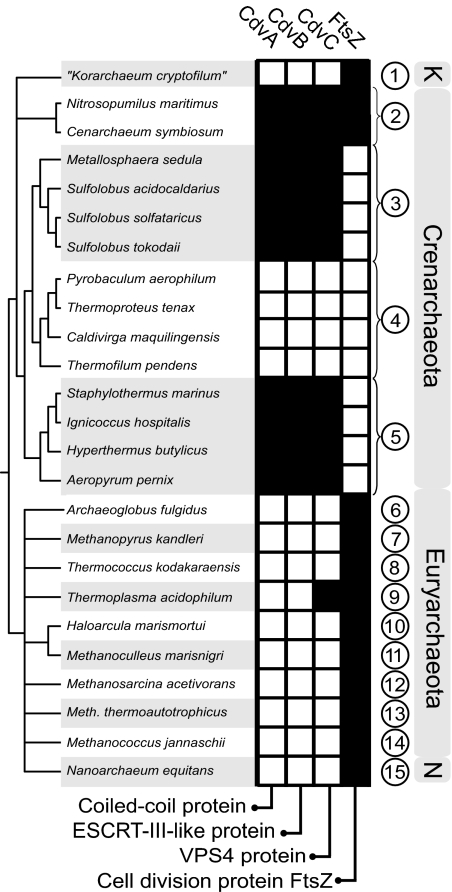
In contrast to the cell division machineries of bacteria, euryarchaea, and eukaryotes, no division components have been identified in the second main archaeal phylum, Crenarchaeota. Here, we demonstrate that a three-gene operon, cdv, in the crenarchaeon Sulfolobus acidocaldarius, forms part of a unique cell division machinery. The operon is induced at the onset of genome segregation and division, and the Cdv proteins then polymerize between segregating nucleoids and persist throughout cell division, forming a successively smaller structure during constriction. The cdv operon is dramatically down-regulated after UV irradiation, indicating division inhibition in response to DNA damage, reminiscent of eukaryotic checkpoint systems. The cdv genes exhibit a complementary phylogenetic range relative to FtsZ-based archaeal division systems such that, in most archaeal lineages, either one or the other system is present. Two of the Cdv proteins, CdvB and CdvC, display homology to components of the eukaryotic ESCRT-III sorting complex involved in budding of luminal vesicles and HIV-1 virion release, suggesting mechanistic similarities and a common evolutionary origin.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Comment in
-
Cell sorting protein homologs reveal an unusual diversity in archaeal cell division.Proc Natl Acad Sci U S A. 2008 Dec 2;105(48):18653-4. doi: 10.1073/pnas.0810505106. Epub 2008 Nov 24. Proc Natl Acad Sci U S A. 2008. PMID: 19033202 Free PMC article. No abstract available.
References
-
- Lundgren M, Bernander R. Archaeal cell cycle progress. Curr Opin Microbiol. 2005;8:662–668. - PubMed
-
- Bernander R. The cell cycle of Sulfolobus. Mol Microbiol. 2007;66:557–562. - PubMed
-
- Morgan DO. In: The Cell Cycle: Principles of Control. Lawrence E, editor. Sunderland: New Science Press Ltd.; 2007. pp. 157–173.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
