

Strain-specific differences in the mechanisms of progesterone regulation of murine mammary gland development
- PMID: 18988671
- PMCID: PMC2654739
- DOI: 10.1210/en.2008-1459
Strain-specific differences in the mechanisms of progesterone regulation of murine mammary gland development
Abstract
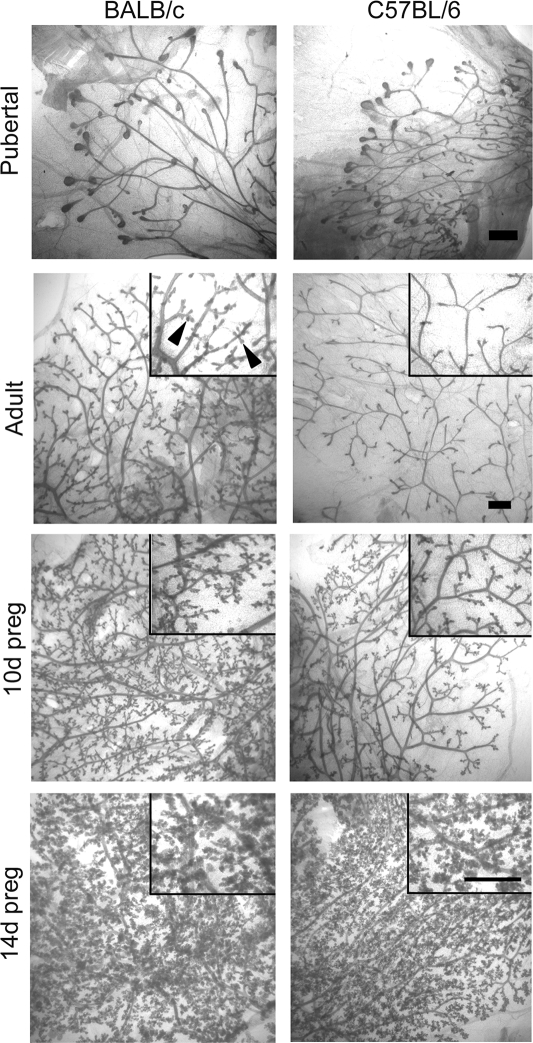
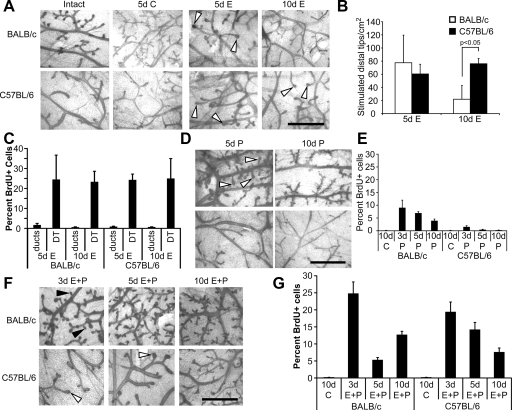
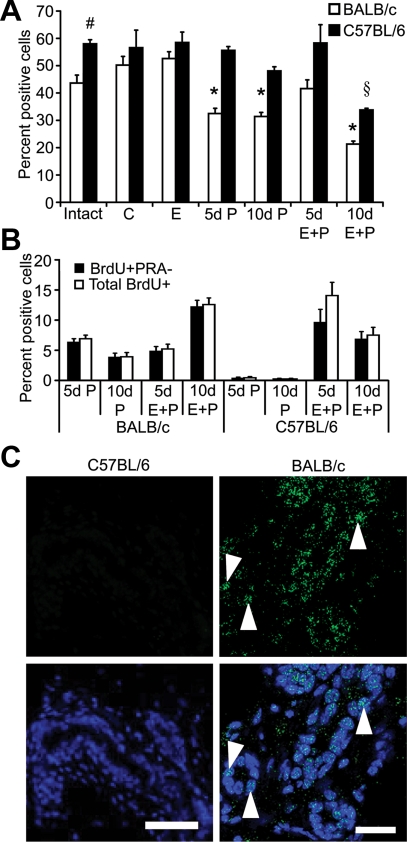
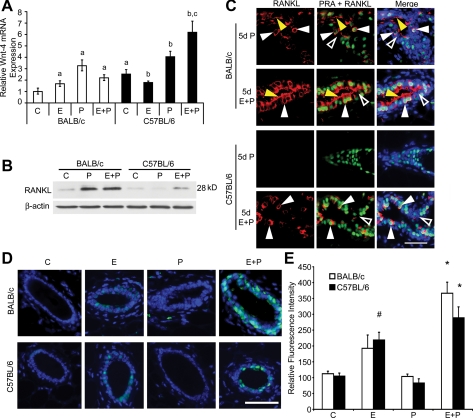
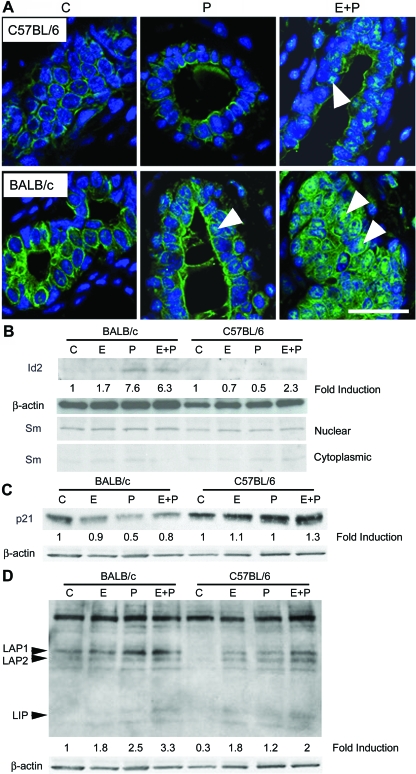
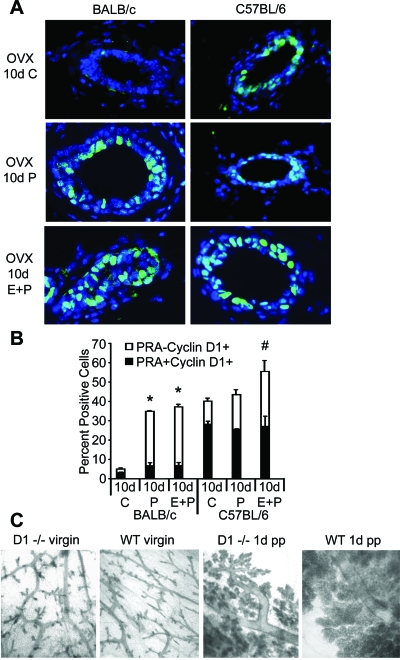
Progesterone (P) is required for normal mammary gland development, and is implicated in the etiology of mammary cancer in rodents and humans. We analyzed mammary gland developmental responses to P and estrogen (E) in two strains of mice (BALB/c and C57BL/6) that exhibit differences in ductal development at sexual maturity and alveologenesis during pregnancy. C57BL/6 mice exhibited reduced proliferative and morphological responses to P. Analysis of known mediators of sidebranching and alveologenesis revealed that reduced P-induced expression of P receptor isoform B and receptor activator of nuclear factor-kappaB ligand (RANKL), as well as altered expression and regulation of cyclin D1, CCAAT/enhancer binding protein beta, and the downstream effectors of RANKL, nuclear Id2 and p21, contribute significantly to the reduced P responsiveness of the C57BL/6 mammary gland. In contrast, E responsiveness was greater in C57BL/6 than in BALB/c glands. E may play a compensatory role in C57BL/6 alveologenesis through its effect on the induction and activation of signal transducer and activator of transcription 5a, a known regulator of RANKL. These observations suggest that in human populations with heterogeneous genetic backgrounds, individuals may respond differentially to the same hormone. Thus, genetic diversity may have a role in determining the effects of P in normal mammary development and tumorigenesis.
Figures
References
-
- Aupperlee MD, Haslam SZ 2007 Differential hormonal regulation and function of progesterone receptor isoforms in normal adult mouse mammary gland. Endocrinology 148:2290–2300 - PubMed
-
- Fendrick JL, Raafat AM, Haslam SZ 1998 Mammary gland growth and development from the postnatal period to postmenopause: ovarian steroid receptor ontogeny and regulation in the mouse. J Mammary Gland Biol Neoplasia 3:7–22 - PubMed
-
- Hofseth LJ, Raafat AM, Osuch JR, Pathak DR, Slomski CA, Haslam SZ 1999 Hormone replacement therapy with estrogen or estrogen plus medroxyprogesterone acetate is associated with increased epithelial proliferation in the normal postmenopausal breast. J Clin Endocrinol Metab 84:4559–4565 - PubMed
-
- Graham JD, Yager ML, Hill HD, Byth K, O'Neill GM, Clarke CL 2005 Altered progesterone receptor isoform expression remodels progestin responsiveness of breast cancer cells. Mol Endocrinol 19:2713–2735 - PubMed
-
- Graham JD, Yeates C, Balleine RL, Harvey SS, Milliken JS, Bilous AM, Clarke CL 1996 Progesterone receptor A and B protein expression in human breast cancer. J Steroid Biochem Mol Biol 56:93–98 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
