

Cytosolic chaperones influence the fate of a toxin dislocated from the endoplasmic reticulum
- PMID: 18988734
- PMCID: PMC2580750
- DOI: 10.1073/pnas.0809013105
Cytosolic chaperones influence the fate of a toxin dislocated from the endoplasmic reticulum
Abstract
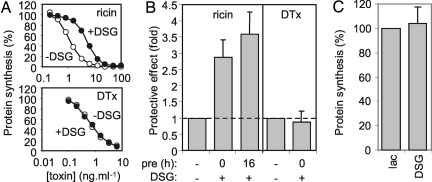
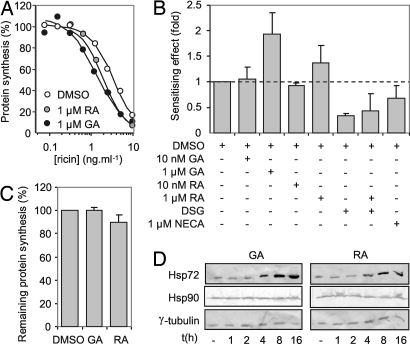
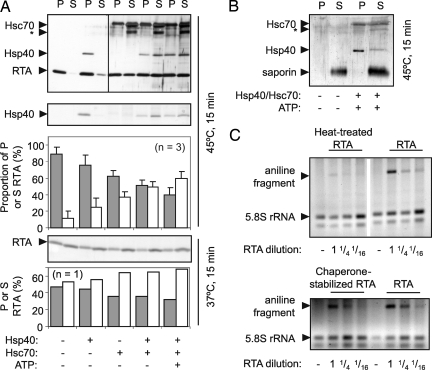
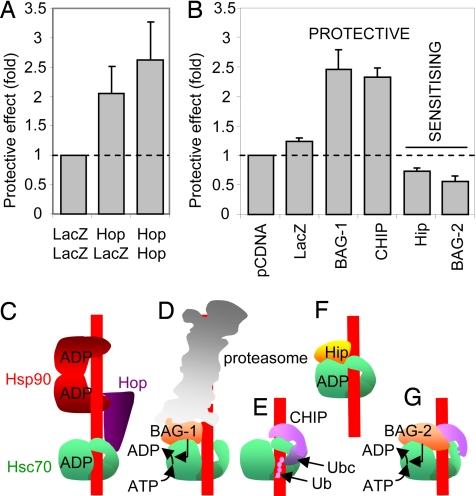
The plant cytotoxin ricin enters target mammalian cells by receptor-mediated endocytosis and undergoes retrograde transport to the endoplasmic reticulum (ER). Here, its catalytic A chain (RTA) is reductively separated from the cell-binding B chain, and free RTA enters the cytosol where it inactivates ribosomes. Cytosolic entry requires unfolding of RTA and dislocation across the ER membrane such that it arrives in the cytosol in a vulnerable, nonnative conformation. Clearly, for such a dislocated toxin to become active, it must avoid degradation and fold to a catalytic conformation. Here, we show that, in vitro, Hsc70 prevents aggregation of heat-treated RTA, and that RTA catalytic activity is recovered after chaperone treatment. A combination of pharmacological inhibition and cochaperone expression reveals that, in vivo, cytosolic RTA is scrutinized sequentially by the Hsc70 and Hsp90 cytosolic chaperone machineries, and that its eventual fate is determined by the balance of activities of cochaperones that regulate Hsc70 and Hsp90 functions. Cytotoxic activity follows Hsc70-mediated escape of RTA from an otherwise destructive pathway facilitated by Hsp90. We demonstrate a role for cytosolic chaperones, proteins typically associated with folding nascent proteins, assembling multimolecular protein complexes and degrading cytosolic and stalled, cotranslocational clients, in a toxin triage, in which both toxin folding and degradation are initiated from chaperone-bound states.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Meusser B, Hirsch C, Jarosch E, Sommer T. ERAD: The long road to destruction. Nat Cell Biol. 2005;7:766–772. - PubMed
-
- Wesche J, Rapak A, Olsnes S. Dependence of ricin toxicity on translocation of the toxin A-chain from the endoplasmic reticulum to the cytosol. J Biol Chem. 1999;274:34443–34449. - PubMed
-
- Argent RH, et al. Introduction of a disulfide bond into ricin A chain decreases the cytotoxicity of the ricin holotoxin. J Biol Chem. 1994;269:26705–26710. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
