

Fat metabolism links germline stem cells and longevity in C. elegans
- PMID: 18988854
- PMCID: PMC2760269
- DOI: 10.1126/science.1162011
Fat metabolism links germline stem cells and longevity in C. elegans
Abstract
Fat metabolism, reproduction, and aging are intertwined regulatory axes; however, the mechanism by which they are coupled remains poorly understood. We found that germline stem cells (GSCs) actively modulate lipid hydrolysis in Caenorhabditis elegans, which in turn regulates longevity. GSC arrest promotes systemic lipolysis via induction of a specific fat lipase. Subsequently, fat mobilization is promoted and life span is prolonged. Constitutive expression of this lipase in fat storage tissue generates lean and long-lived animals. This lipase is a key factor in the lipid hydrolysis and increased longevity that are induced by decreased insulin signaling. These results suggest a link between C. elegans fat metabolism and longevity.
Figures
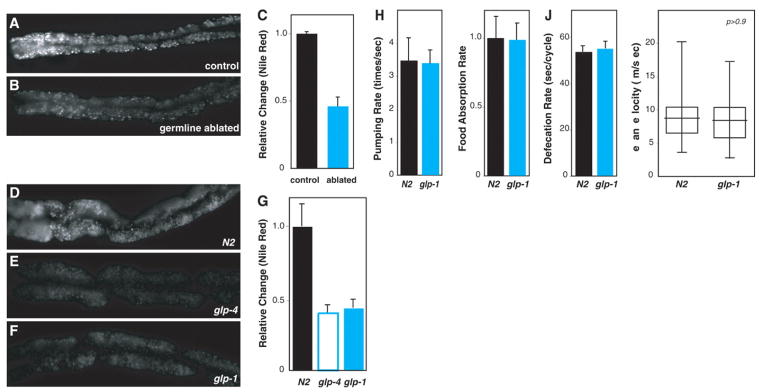
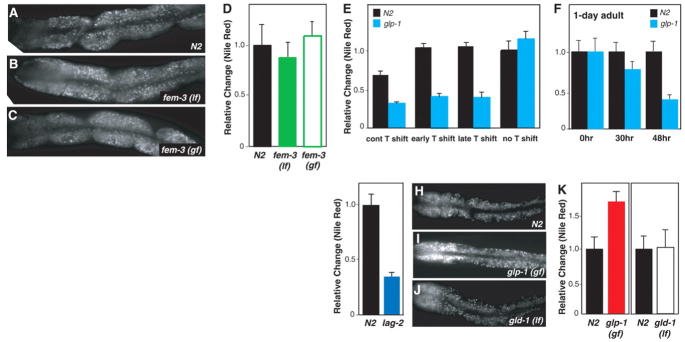
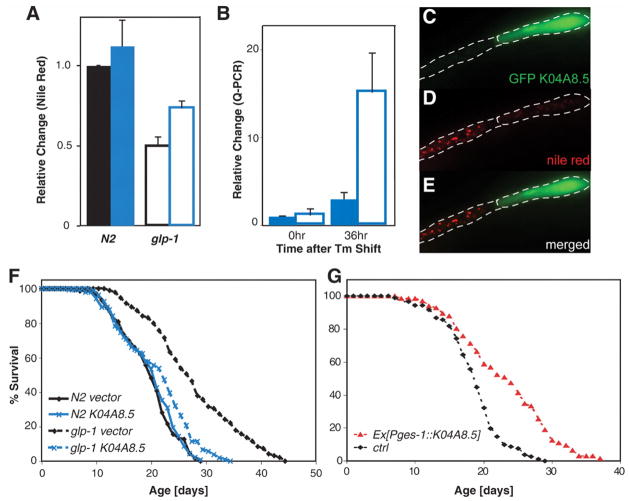
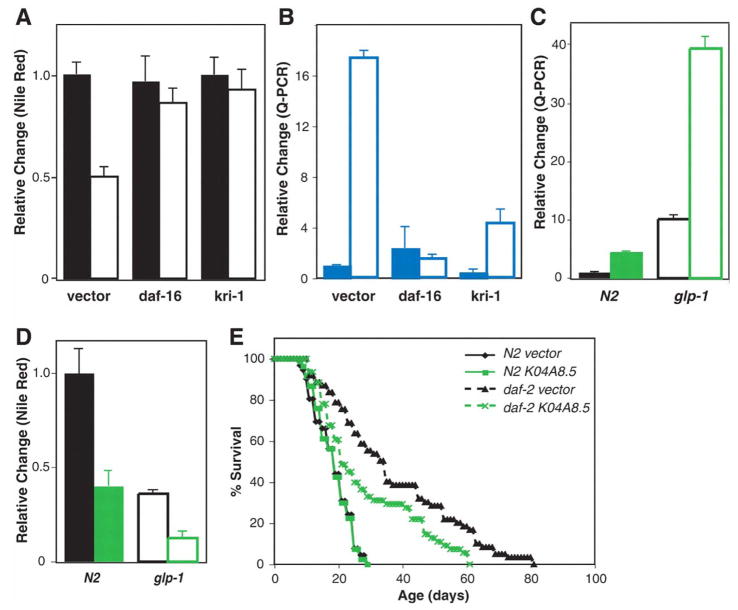
Comment in
-
Physiology. Burn fat, live longer.Science. 2008 Nov 7;322(5903):865-6. doi: 10.1126/science.1166150. Science. 2008. PMID: 18988829 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
