

Acute internalization of gap junctions in vascular endothelial cells in response to inflammatory mediator-induced G-protein coupled receptor activation
- PMID: 18992245
- PMCID: PMC2628571
- DOI: 10.1016/j.febslet.2008.10.043
Acute internalization of gap junctions in vascular endothelial cells in response to inflammatory mediator-induced G-protein coupled receptor activation
Abstract
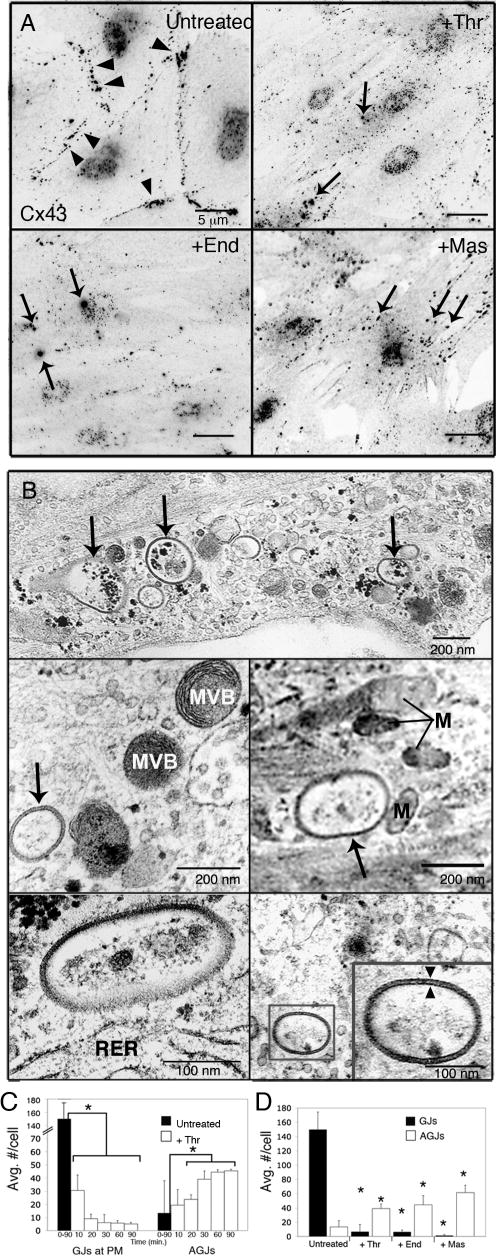
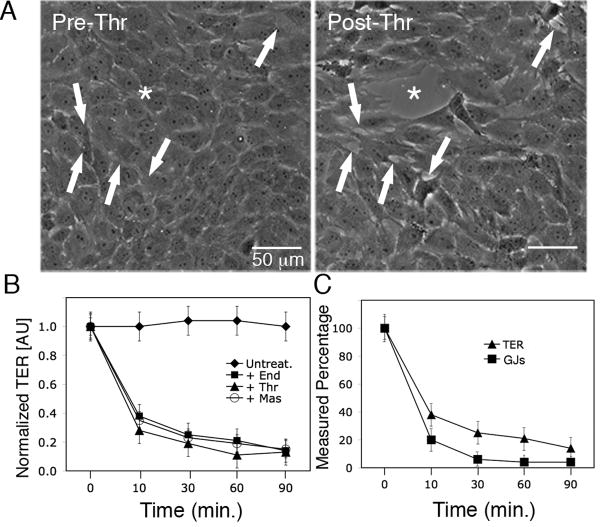
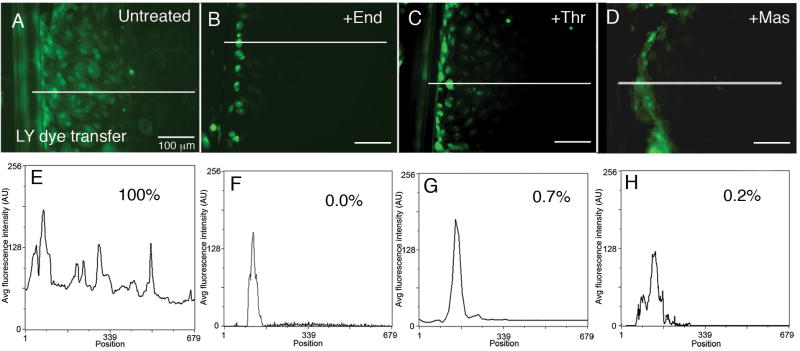
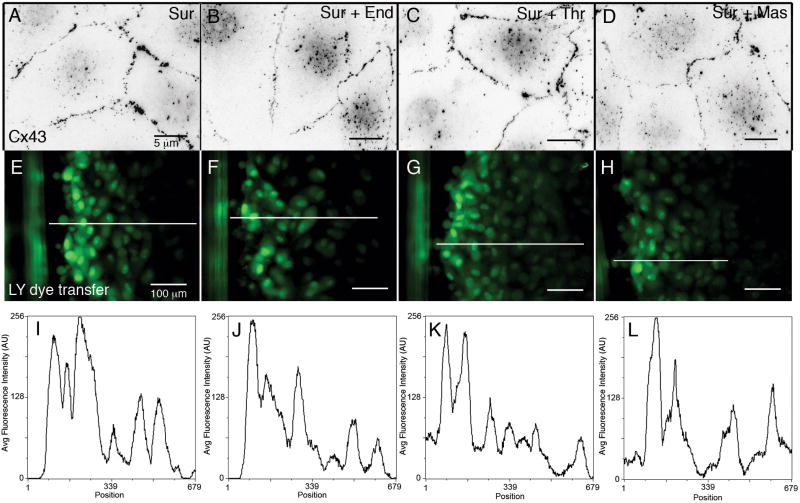
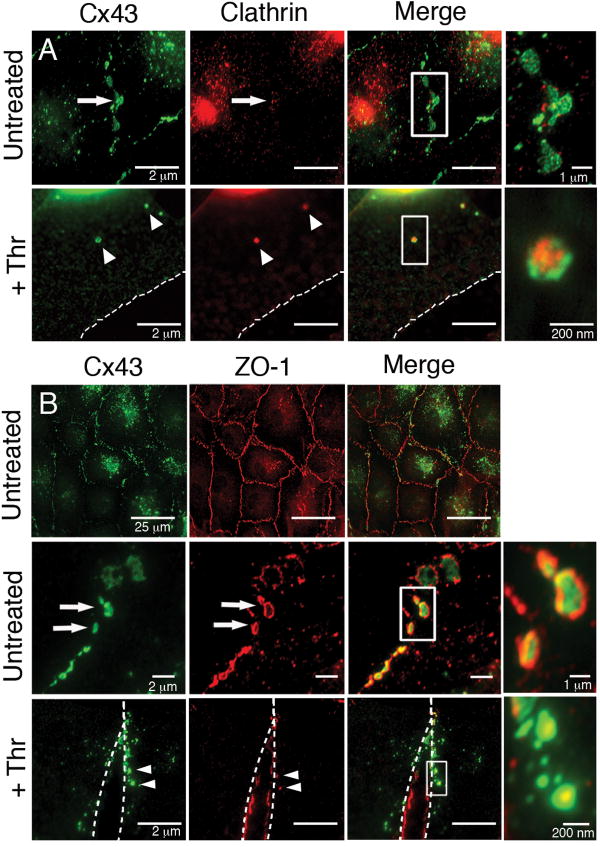
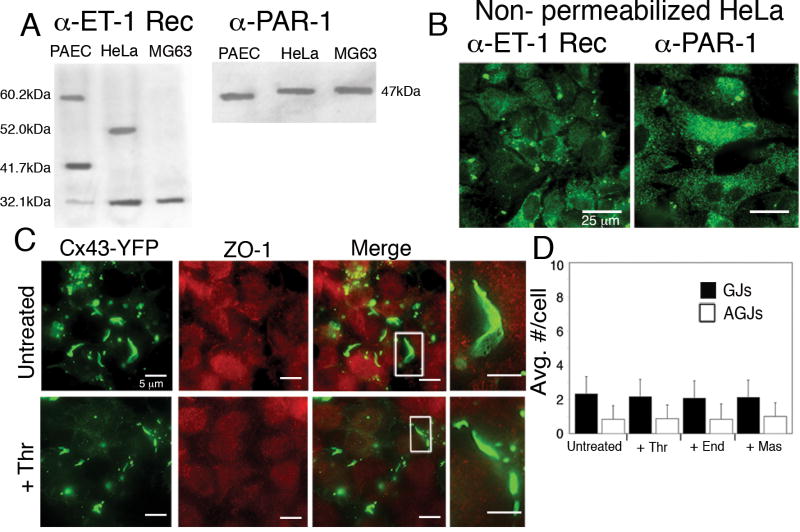
During the inflammatory response, activation of G-protein coupled receptors (GPCRs) by inflammatory mediators rapidly leads to inhibition of gap junction intercellular communication (GJIC); however, the steps that lead to this inhibition are not known. Combining high-resolution fluorescence microscopy and functional assays, we found that activation of the GPCRs PAR-1 and ET(A/B) by their natural inflammatory mediator agonists, thrombin and endothelin-1, resulted in rapid and acute internalization of gap junctions (GJs) that coincided with the inhibition of GJIC followed by increased vascular permeability. The endocytosis protein clathrin and the scaffold protein ZO-1 appeared to be involved in GJ internalization, and ZO-1 was partially displaced from GJs during the internalization process. These findings demonstrate that GJ internalization is an efficient mechanism for modulating GJIC in inflammatory response.
Figures
References
-
- Mehta PP, Bertram JS, Loewenstein WR. Growth inhibition of transformed cells correlates with their junctional communication with normal cells. Cell. 1986;44:187–96. - PubMed
-
- Loewenstein WR, Rose B. The cell-cell channel in the control of growth. Semin Cell Biol. 1992;3:59–79. - PubMed
-
- Guthrie SC, Gilula NB. Gap junctional communication and development. Trends Neurosci. 1989;12:12–6. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
