

Receptor-independent, direct membrane binding leads to cell-surface lipid sorting and Syk kinase activation in dendritic cells
- PMID: 18993083
- PMCID: PMC2642965
- DOI: 10.1016/j.immuni.2008.09.013
Receptor-independent, direct membrane binding leads to cell-surface lipid sorting and Syk kinase activation in dendritic cells
Abstract
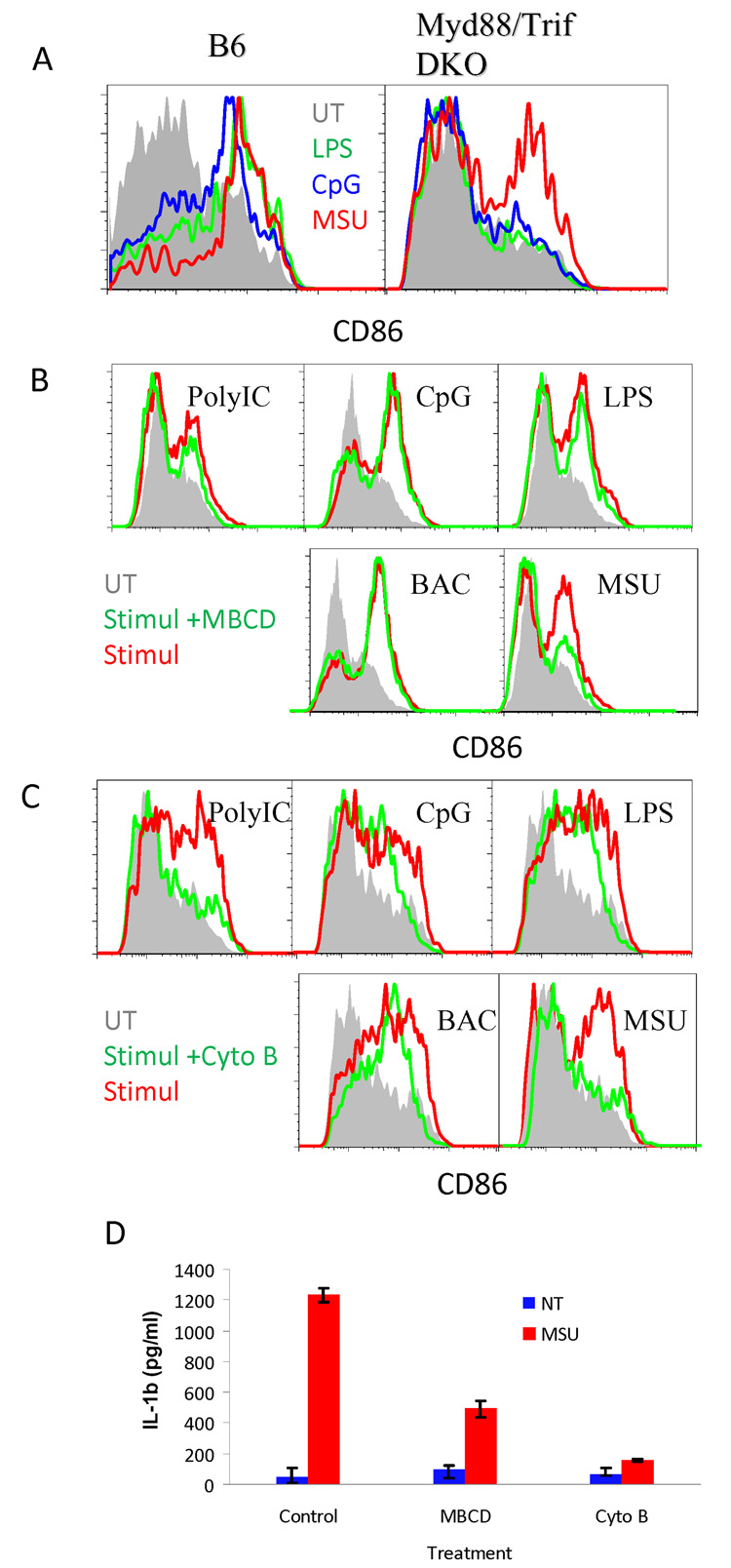
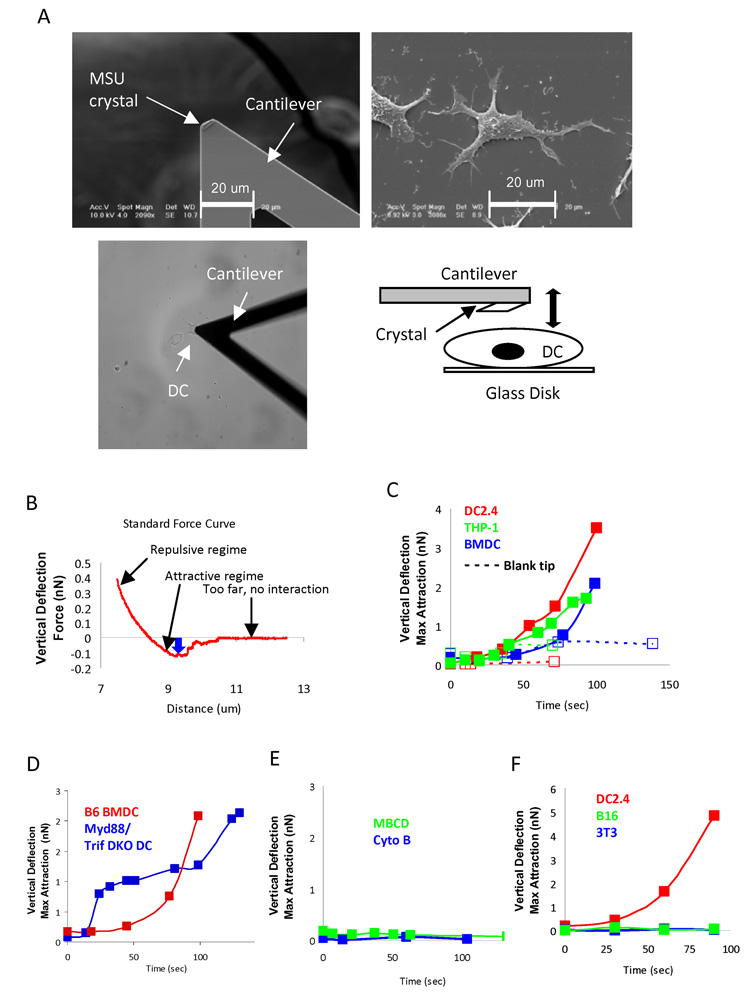
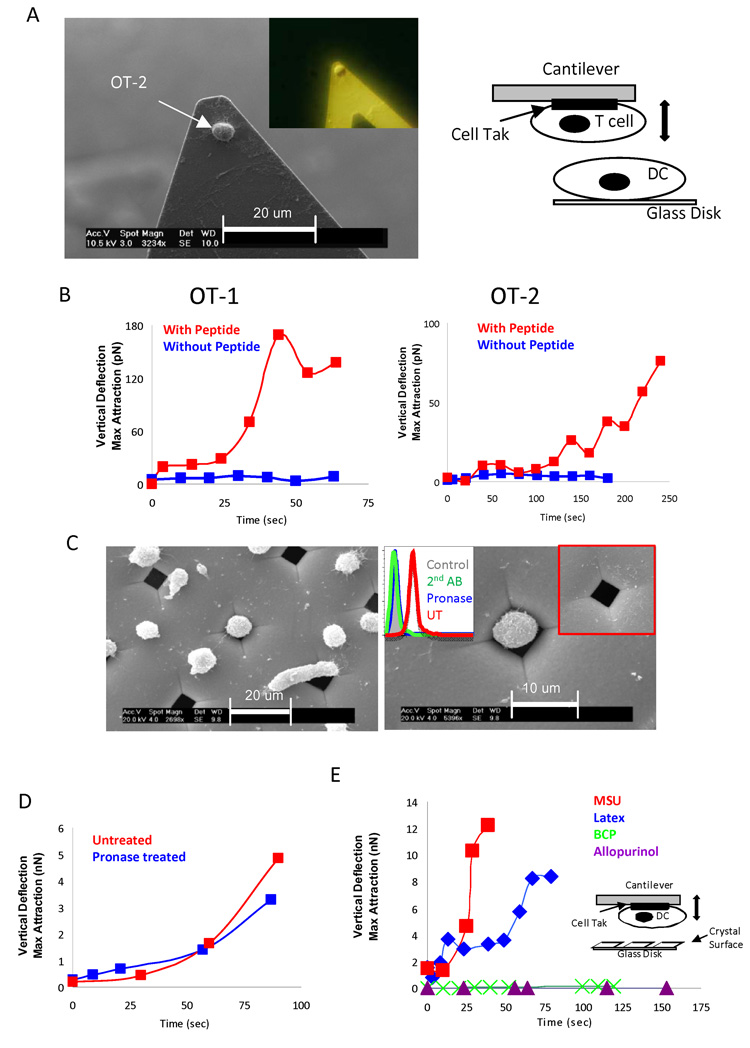
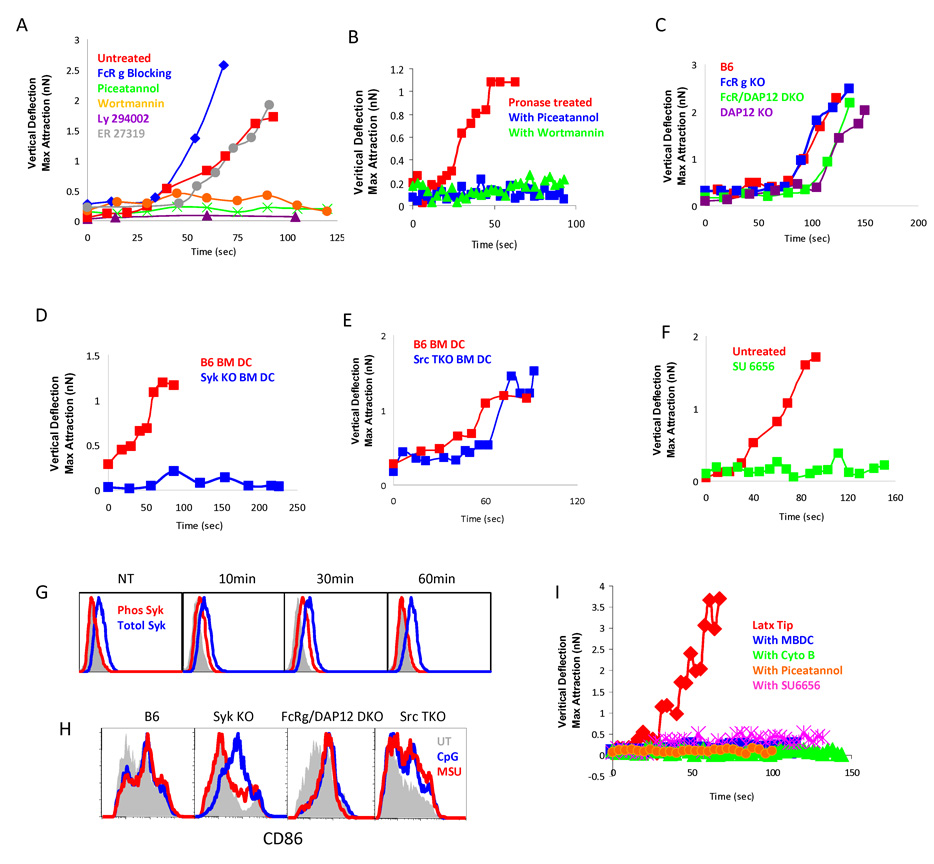
Binding of particulate antigens by antigen-presenting cells is a critical step in immune activation. Previously, we demonstrated that uric acid crystals are potent adjuvants, initiating a robust adaptive immune response. However, the mechanisms of activation are unknown. By using atomic force microscopy as a tool for real-time single-cell activation analysis, we report that uric acid crystals could directly engage cellular membranes, particularly the cholesterol components, with a force substantially stronger than protein-based cellular contacts. Binding of particulate substances activated Syk kinase-dependent signaling in dendritic cells. These observations suggest a mechanism whereby immune cell activation can be triggered by solid structures via membrane lipid alteration without the requirement for specific cell-surface receptors, and a testable hypothesis for crystal-associated arthropathies, inflammation, and adjuvanticity.
Figures
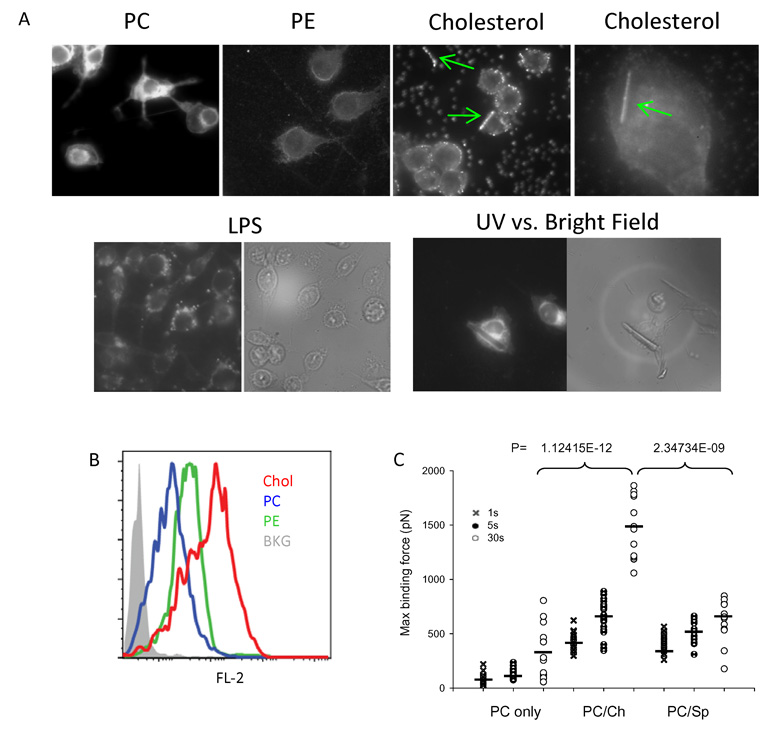
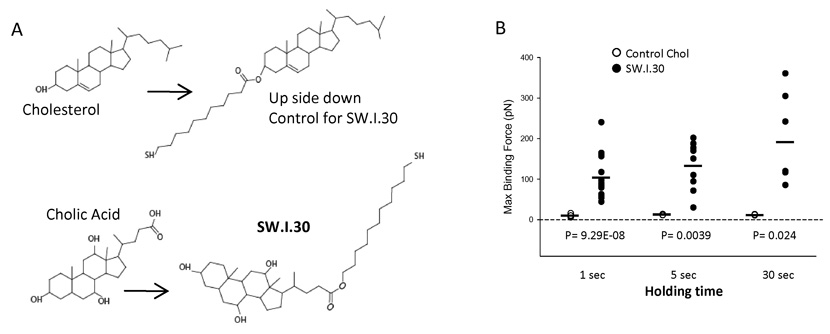
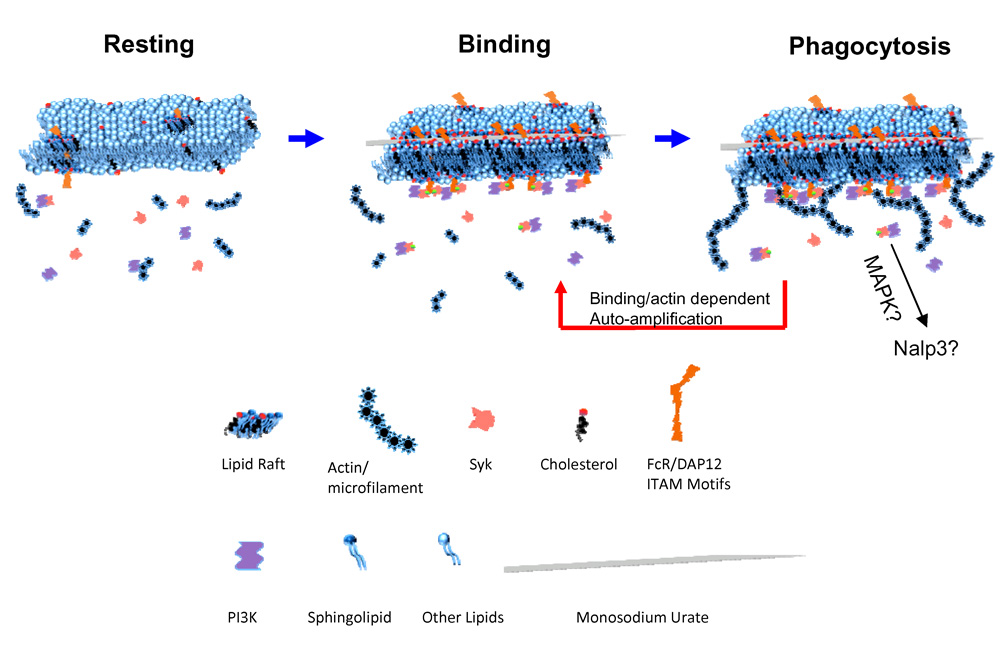
Comment in
-
"Dangerous crystals".Immunity. 2008 Nov 14;29(5):670-1. doi: 10.1016/j.immuni.2008.10.005. Immunity. 2008. PMID: 19006690
References
-
- Abram CL, Lowell CA. The expanding role for ITAM-based signaling pathways in immune cells. Sci STKE. 2007:re2. - PubMed
-
- Aderem A, Underhill DM. Mechanisms of phagocytosis in macrophages. Annu Rev Immunol. 1999;17:593–623. - PubMed
-
- Brown DA. Lipid rafts, detergent-resistant membranes, and raft targeting signals. Physiology (Bethesda) 2006;21:430–439. - PubMed
-
- Brown DA, London E. Functions of lipid rafts in biological membranes. Annu Rev Cell Dev Biol. 1998;14:111–136. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
