

Mucosal glycan foraging enhances fitness and transmission of a saccharolytic human gut bacterial symbiont
- PMID: 18996345
- PMCID: PMC2605320
- DOI: 10.1016/j.chom.2008.09.007
Mucosal glycan foraging enhances fitness and transmission of a saccharolytic human gut bacterial symbiont
Abstract
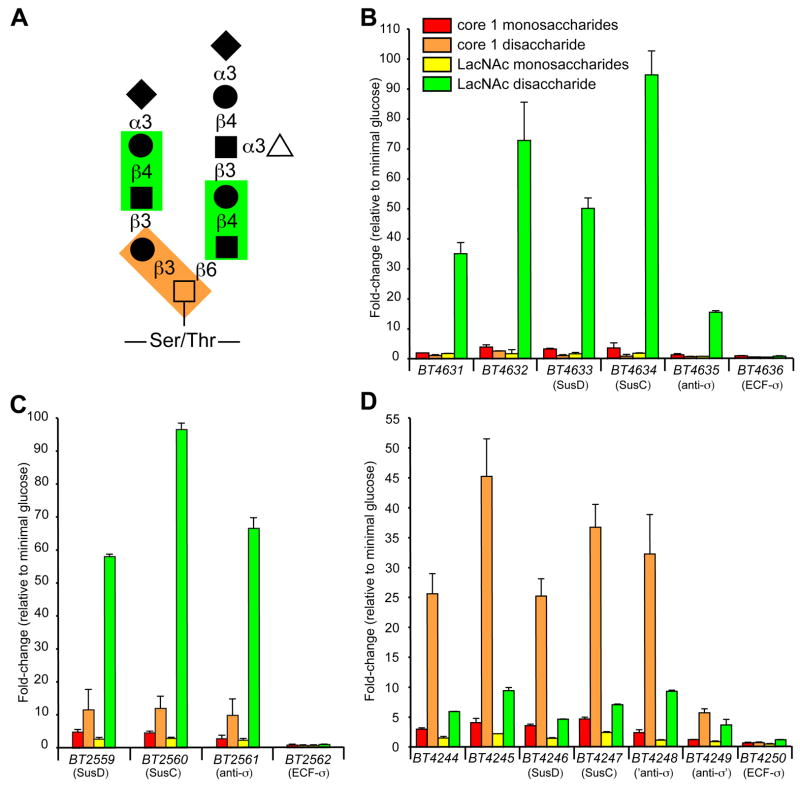
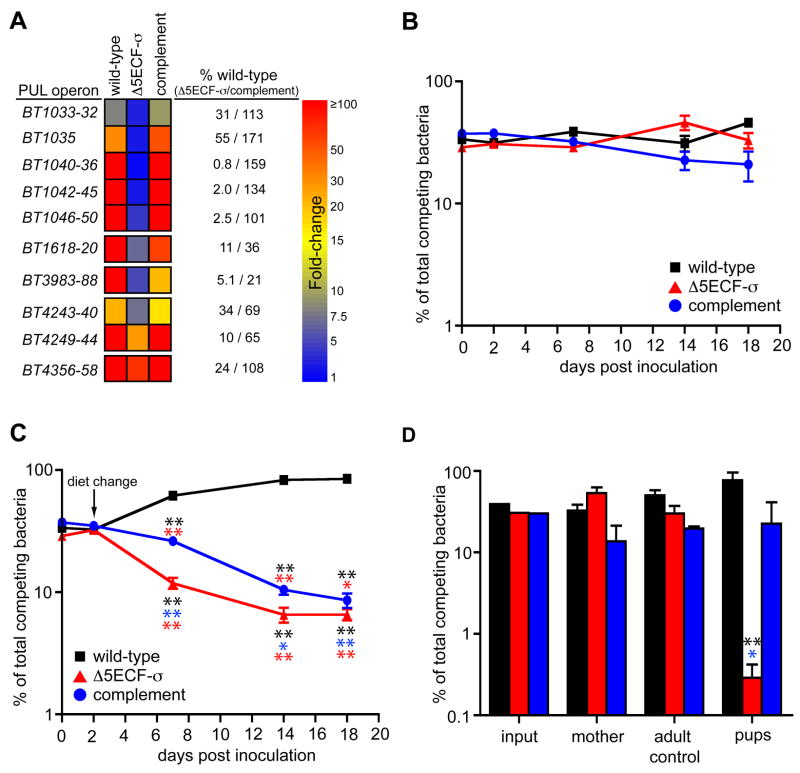
The distal human gut is a microbial bioreactor that digests complex carbohydrates. The strategies evolved by gut microbes to sense and process diverse glycans have important implications for the assembly and operation of this ecosystem. The human gut-derived bacterium Bacteroides thetaiotaomicron forages on both host and dietary glycans. Its ability to target these substrates resides in 88 polysaccharide utilization loci (PULs), encompassing 18% of its genome. Whole genome transcriptional profiling and genetic tests were used to define the mechanisms underlying host glycan foraging in vivo and in vitro. PULs that target all major classes of host glycans were identified. However, mucin O-glycans are the principal host substrate foraged in vivo. Simultaneous deletion of five genes encoding ECF-sigma transcription factors, which activate mucin O-glycan utilization, produces defects in bacterial persistence in the gut and in mother-to-offspring transmission. Thus, PUL-mediated glycan catabolism is an important component in gut colonization and may impact microbiota ecology.
Figures
References
-
- Bayer EA, Belaich JP, Shoham Y, Lamed R. The cellulosomes: multienzyme machines for degradation of plant cell wall polysaccharides. Ann Rev Microbiol. 2004;58:521–554. - PubMed
-
- Bjursell MK, Martens EC, Gordon JI. Functional genomic and metabolic studies of the adaptations of a prominent adult human gut symbiont, Bacteroides thetaiotaomicron, to the suckling period. J Biol Chem. 2006;281:36269–36279. - PubMed
-
- Braun V, Mahren S. Transmembrane transcriptional control (surface signalling) of the Escherichia coli Fec type. FEMS Microbiol Rev. 2005;29:673–684. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 DK030292/DK/NIDDK NIH HHS/United States
- R01 DK070977/DK/NIDDK NIH HHS/United States
- F32 AI073060/AI/NIAID NIH HHS/United States
- P30 DK056341/DK/NIDDK NIH HHS/United States
- T32 HD07409/HD/NICHD NIH HHS/United States
- T32 HG000045/HG/NHGRI NIH HHS/United States
- K01 DK084214/DK/NIDDK NIH HHS/United States
- R37 DK030292/DK/NIDDK NIH HHS/United States
- K22 HG000045/HG/NHGRI NIH HHS/United States
- HG00045/HG/NHGRI NIH HHS/United States
- T32 GM007200/GM/NIGMS NIH HHS/United States
- DK30292/DK/NIDDK NIH HHS/United States
- GM07200/GM/NIGMS NIH HHS/United States
- T32 HD007409/HD/NICHD NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
