

Propagating waves of activity in the neocortex: what they are, what they do
- PMID: 18997124
- PMCID: PMC2679998
- DOI: 10.1177/1073858408317066
Propagating waves of activity in the neocortex: what they are, what they do
Abstract
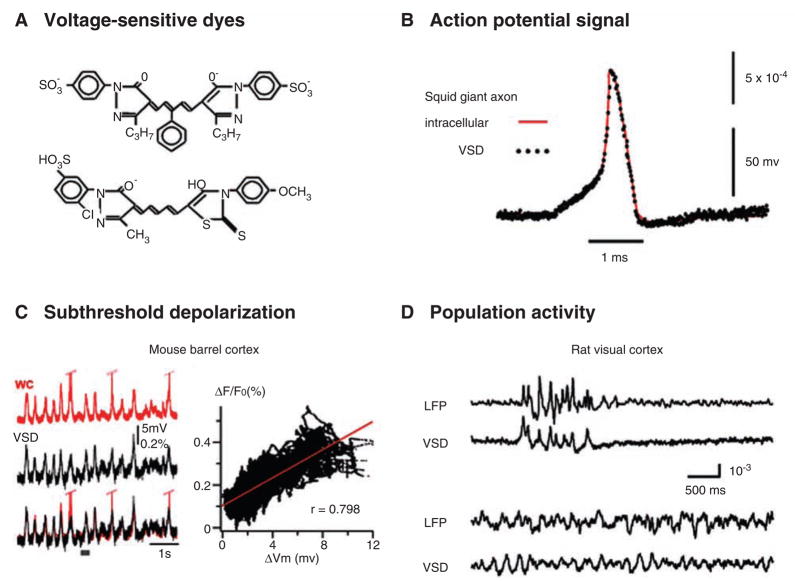
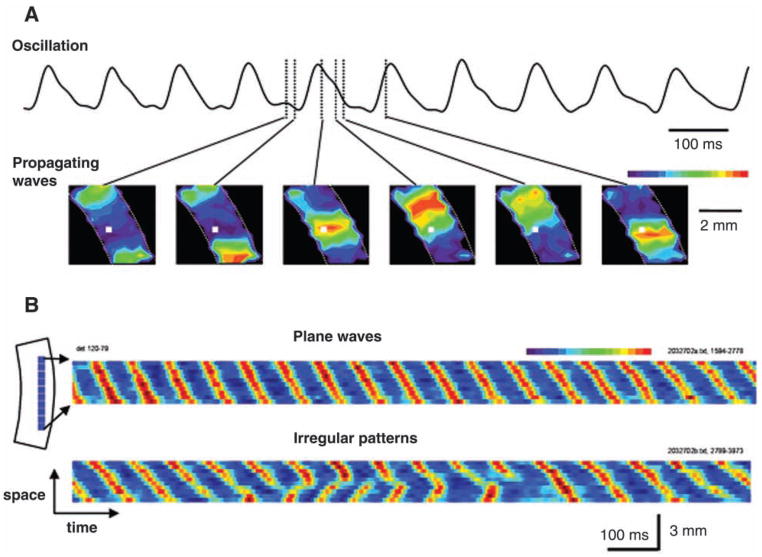
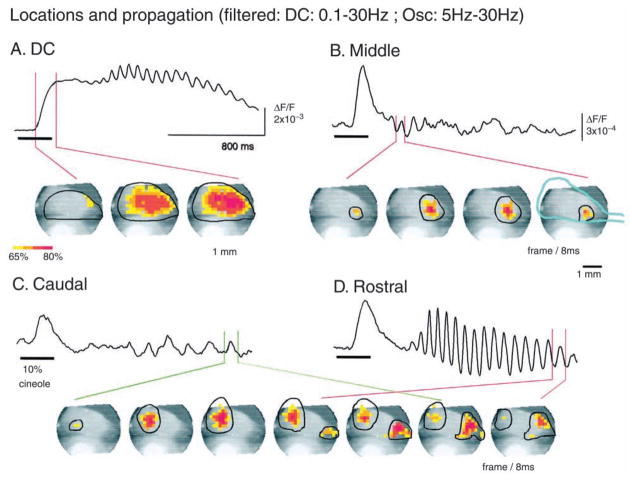
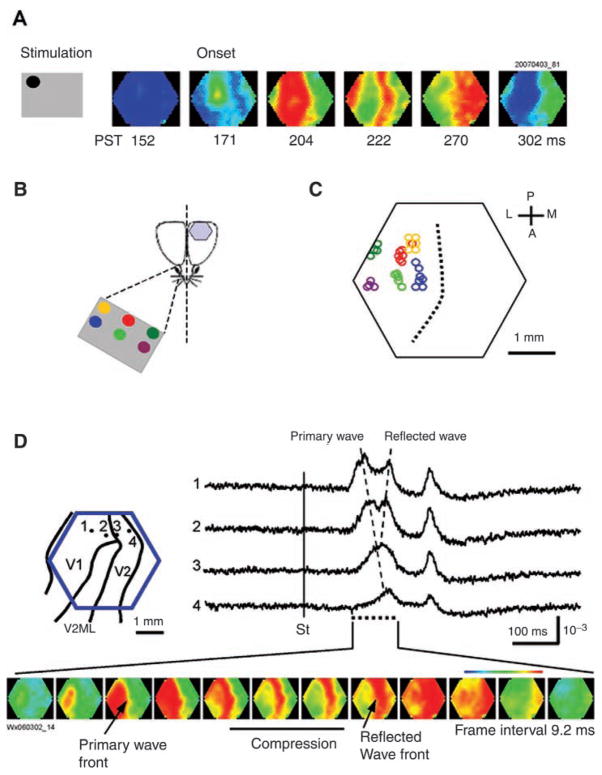
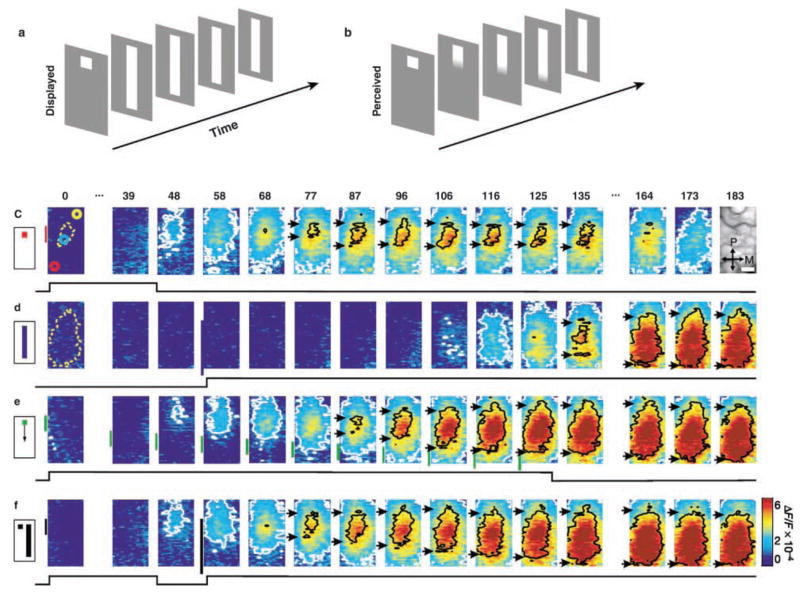
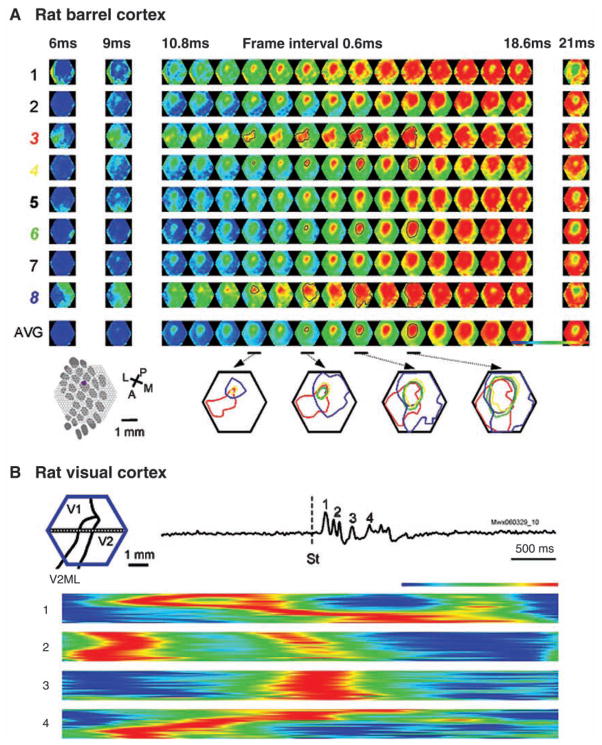
The development of voltage-sensitive dyes (VSD) and fast optical imaging techniques have brought us a new tool for examining spatiotemporal patterns of population neuronal activity in the neocortex. Propagating waves have been observed during almost every type of cortical processing examined by VSD imaging or electrode arrays. These waves provide subthreshold depolarization to individual neurons and increase their spiking probability. Therefore, the propagation of the waves sets up a spatiotemporal framework for increased excitability in neuronal populations, which can help to determine when and where the neurons are likely to fire. In this review, first discussed is propagating waves observed in various systems and possible mechanisms for generating and sustaining these waves. Then discussed are wave dynamics as an emergent behavior of the population activity that can, in turn, influence the activity of individual neurons. The functions of spontaneous and sensory-evoked waves remain to be explored. An important next step will be to examine the interaction between dynamics of propagating waves and functions in the cortex, and to verify if cortical processing can be modified when these waves are altered.
Figures


Similar articles
-
Reverberation of recent visual experience in spontaneous cortical waves.Neuron. 2008 Oct 23;60(2):321-7. doi: 10.1016/j.neuron.2008.08.026. Neuron. 2008. PMID: 18957223 Free PMC article.
-
Columnar interactions determine horizontal propagation of recurrent network activity in neocortex.J Neurosci. 2012 Apr 18;32(16):5454-71. doi: 10.1523/JNEUROSCI.5006-11.2012. J Neurosci. 2012. PMID: 22514308 Free PMC article.
-
Origin of the earliest correlated neuronal activity in the chick embryo revealed by optical imaging with voltage-sensitive dyes.Eur J Neurosci. 2009 Jan;29(1):1-13. doi: 10.1111/j.1460-9568.2008.06568.x. Epub 2008 Dec 11. Eur J Neurosci. 2009. PMID: 19077122
-
Neuronal circuits of the neocortex.Annu Rev Neurosci. 2004;27:419-51. doi: 10.1146/annurev.neuro.27.070203.144152. Annu Rev Neurosci. 2004. PMID: 15217339 Review.
-
Molecular logic of neocortical projection neuron specification, development and diversity.Nat Rev Neurosci. 2013 Nov;14(11):755-69. doi: 10.1038/nrn3586. Epub 2013 Oct 9. Nat Rev Neurosci. 2013. PMID: 24105342 Free PMC article. Review.
Cited by
-
Genetically engineered fluorescent voltage reporters.ACS Chem Neurosci. 2012 Aug 15;3(8):585-92. doi: 10.1021/cn300041b. Epub 2012 Jun 6. ACS Chem Neurosci. 2012. PMID: 22896802 Free PMC article. Review.
-
Wave-processing of long-scale information by neuronal chains.PLoS One. 2013;8(2):e57440. doi: 10.1371/journal.pone.0057440. Epub 2013 Feb 27. PLoS One. 2013. PMID: 23460856 Free PMC article.
-
Sequentially activated discrete modules appear as traveling waves in neuronal measurements with limited spatiotemporal sampling.Elife. 2024 Mar 7;12:RP92254. doi: 10.7554/eLife.92254. Elife. 2024. PMID: 38451063 Free PMC article.
-
Neurophysiological and computational principles of cortical rhythms in cognition.Physiol Rev. 2010 Jul;90(3):1195-268. doi: 10.1152/physrev.00035.2008. Physiol Rev. 2010. PMID: 20664082 Free PMC article. Review.
-
Stress impacts sensory variability through cortical sensory activity motifs.Transl Psychiatry. 2020 Jan 21;10(1):20. doi: 10.1038/s41398-020-0713-1. Transl Psychiatry. 2020. PMID: 32066714 Free PMC article.
References
-
- Albowitz B, Kuhnt U. Epileptiform activity in the guinea-pig neocortical slice spreads preferentially along supragranular layers—recordings with voltage-sensitive dyes. Eur J Neurosci. 1995;7:1273–84. - PubMed
-
- Anderson WW, Lewis DV, Swartzwelder HS, Wilson WA. Magnesium-free medium activates seizure-like events in the rat hippocampal slice. Brain Res. 1986;398:215–9. - PubMed
-
- Arieli A, Shoham D, Hildesheim R, Grinvald A. Coherent spatiotemporal patterns of ongoing activity revealed by real-time optical imaging coupled with single-unit recording in the cat visual cortex. J Neurophysiol. 1995;73:2072–93. - PubMed
-
- Arieli A, Sterkin A, Grinvald A, Aertsen A. Dynamics of ongoing activity: explanation of the large variability in evoked cortical responses. Science. 1996;273:1868–71. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
