

Deletion of vascular endothelial growth factor in myeloid cells accelerates tumorigenesis
- PMID: 18997773
- PMCID: PMC3103772
- DOI: 10.1038/nature07445
Deletion of vascular endothelial growth factor in myeloid cells accelerates tumorigenesis
Abstract
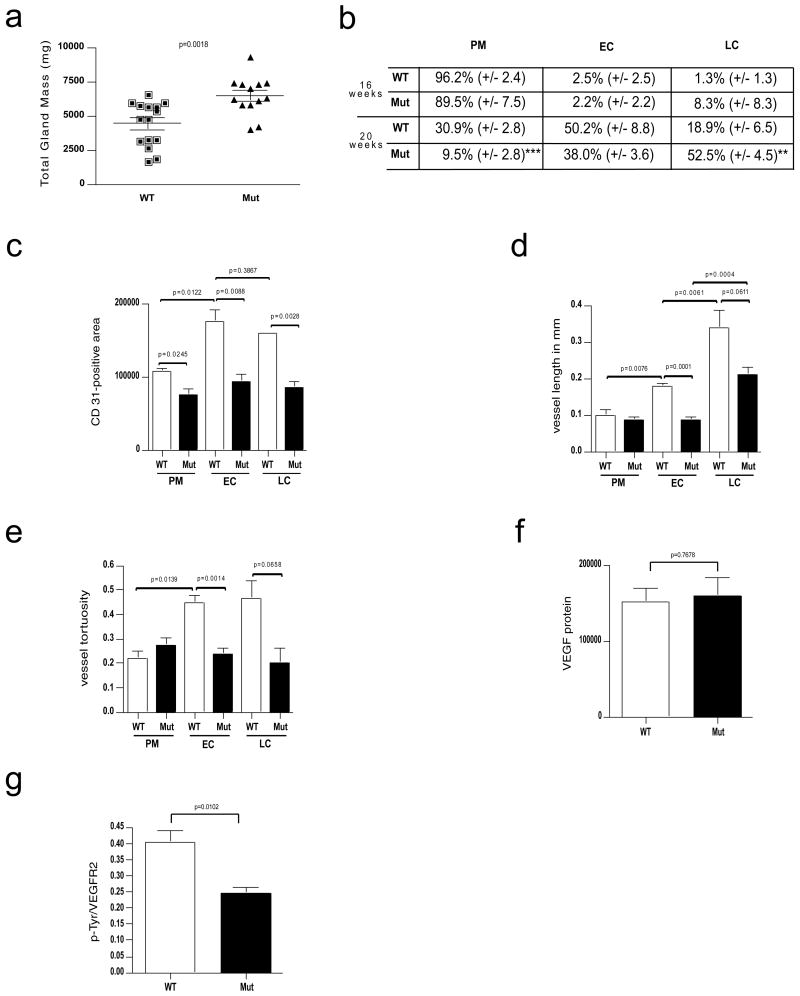
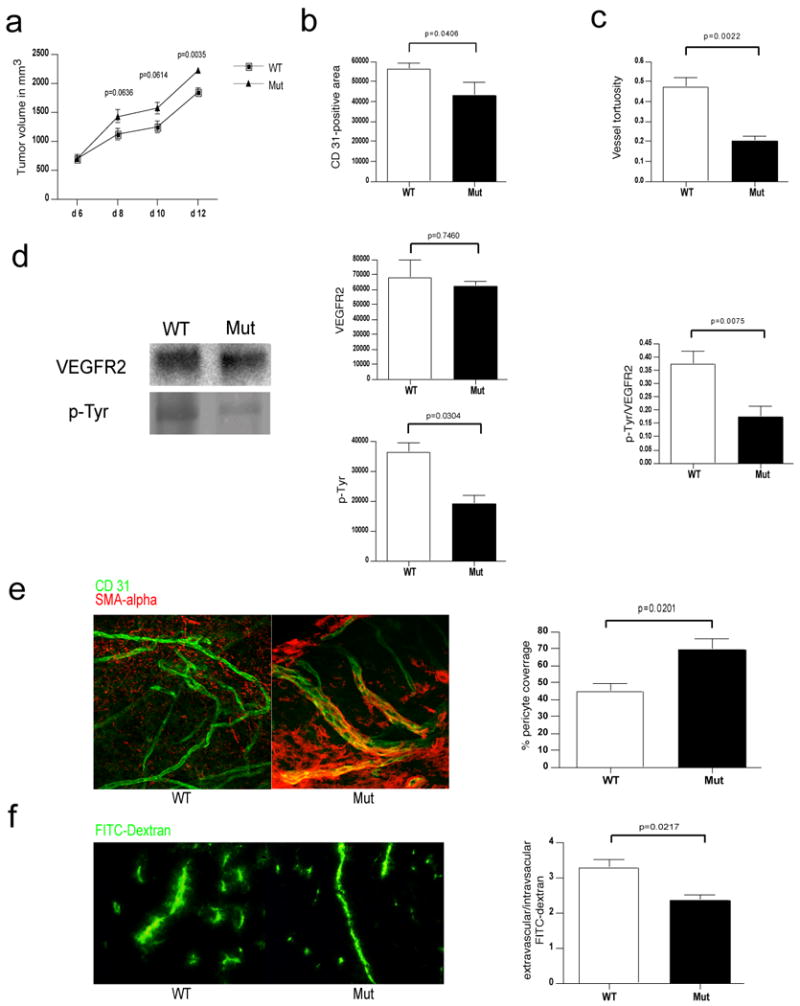
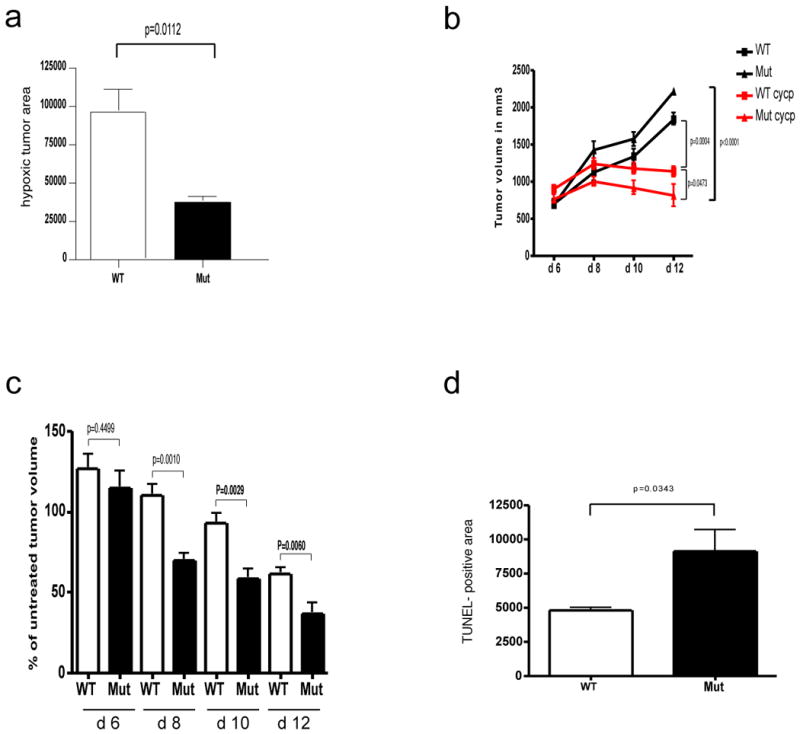
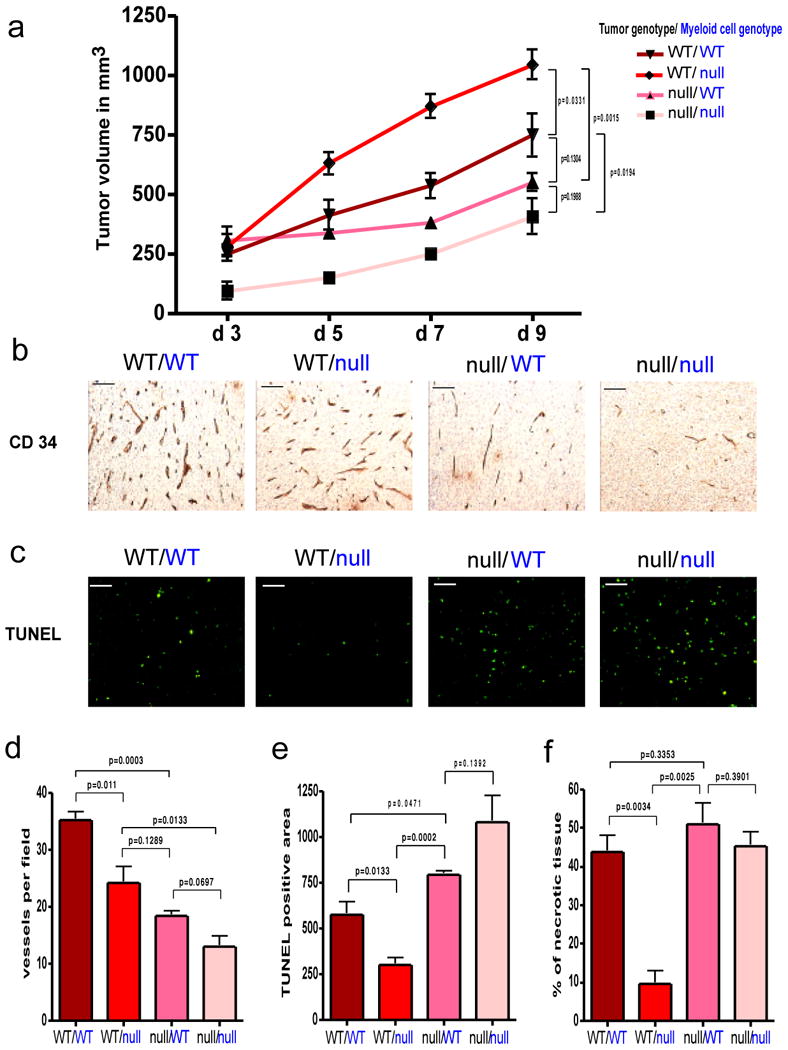
Angiogenesis and the development of a vascular network are required for tumour progression, and they involve the release of angiogenic factors, including vascular endothelial growth factor (VEGF-A), from both malignant and stromal cell types. Infiltration by cells of the myeloid lineage is a hallmark of many tumours, and in many cases the macrophages in these infiltrates express VEGF-A. Here we show that the deletion of inflammatory-cell-derived VEGF-A attenuates the formation of a typical high-density vessel network, thus blocking the angiogenic switch in solid tumours in mice. Vasculature in tumours lacking myeloid-cell-derived VEGF-A was less tortuous, with increased pericyte coverage and decreased vessel length, indicating vascular normalization. In addition, loss of myeloid-derived VEGF-A decreases the phosphorylation of VEGF receptor 2 (VEGFR2) in tumours, even though overall VEGF-A levels in the tumours are unaffected. However, deletion of myeloid-cell VEGF-A resulted in an accelerated tumour progression in multiple subcutaneous isograft models and an autochthonous transgenic model of mammary tumorigenesis, with less overall tumour cell death and decreased tumour hypoxia. Furthermore, loss of myeloid-cell VEGF-A increased the susceptibility of tumours to chemotherapeutic cytotoxicity. This shows that myeloid-derived VEGF-A is essential for the tumorigenic alteration of vasculature and signalling to VEGFR2, and that these changes act to retard, not promote, tumour progression.
Figures
References
-
- Folkman J. Angiogenesis. Annu Rev Med. 2006;57:1–18. - PubMed
-
- Lewis JS, Landers RJ, Underwood JC, Harris AL, Lewis CE. Expression of vascular endothelial growth factor by macrophages is up-regulated in poorly vascularized areas of breast carcinomas. J Pathol. 2000;192(2):150–158. - PubMed
-
- Gerber H, et al. VEGF couples hypertrophic cartilage remodeling, ossification and angiogenesis during endochondral bone formation. Nature Medicine. 1999;5:623–628. - PubMed
-
- Clausen BE, Burkhardt C, Reith W, Renkawitz R, Forster I. Conditional gene targeting in macrophages and granulocytes using LysMcre mice. Transgenic Res. 1999;8(4):265–277. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
