

Thiol-based redox switches in eukaryotic proteins
- PMID: 18999917
- PMCID: PMC2787739
- DOI: 10.1089/ars.2008.2285
Thiol-based redox switches in eukaryotic proteins
Abstract
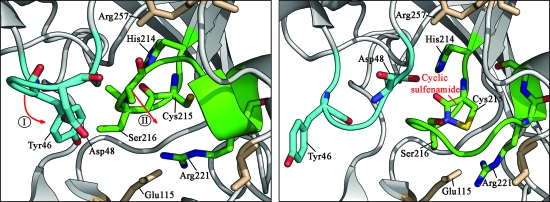
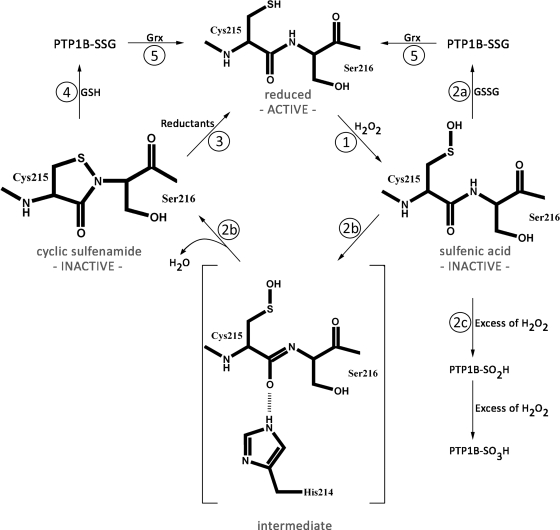
For many years, oxidative thiol modifications in cytosolic proteins were largely disregarded as in vitro artifacts, and considered unlikely to play significant roles within the reducing environment of the cell. Recent developments in in vivo thiol trapping technology combined with mass spectrometric analysis have now provided convincing evidence that thiol-based redox switches are used as molecular tools in many proteins to regulate their activity in response to reactive oxygen and nitrogen species. Reversible oxidative thiol modifications have been found to modulate the function of proteins involved in many different pathways, starting from gene transcription, translation and protein folding, to metabolism, signal transduction, and ultimately apoptosis. This review will focus on three well-characterized eukaryotic proteins that use thiol-based redox switches to influence gene transcription, metabolism, and signal transduction. The transcription factor Yap1p is a good illustration of how oxidative modifications affect the function of a protein without changing its activity. We use glyeraldehyde-3-phosphate dehydrogenase to demonstrate how thiol modification of an active site cysteine re-routes metabolic pathways and converts a metabolic enzyme into a pro-apoptotic factor. Finally, we introduce the redox-sensitive protein tyrosine phosphatase PTP1B to illustrate that reversibility is one of the fundamental aspects of redox-regulation.
Figures
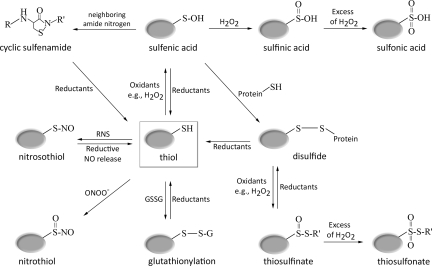
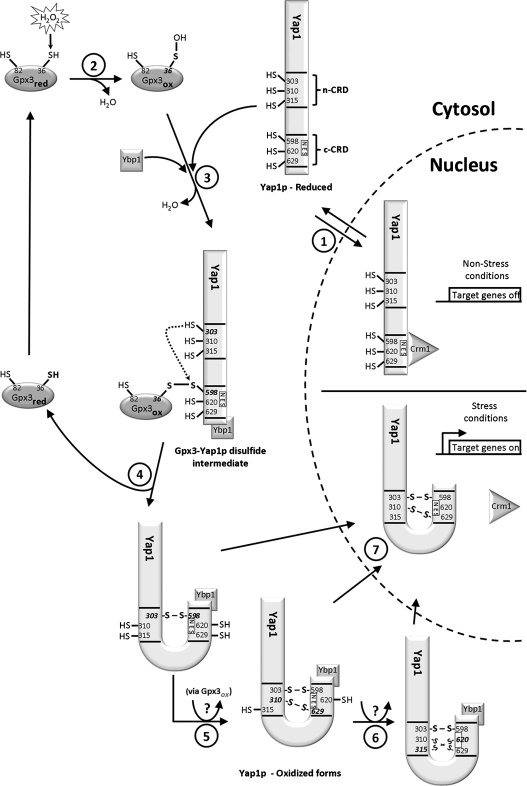
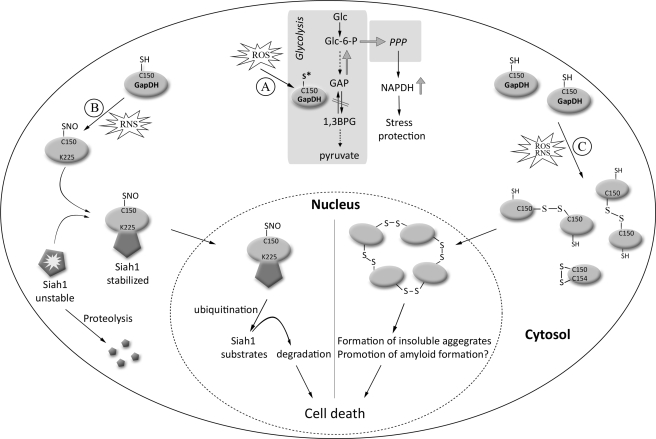
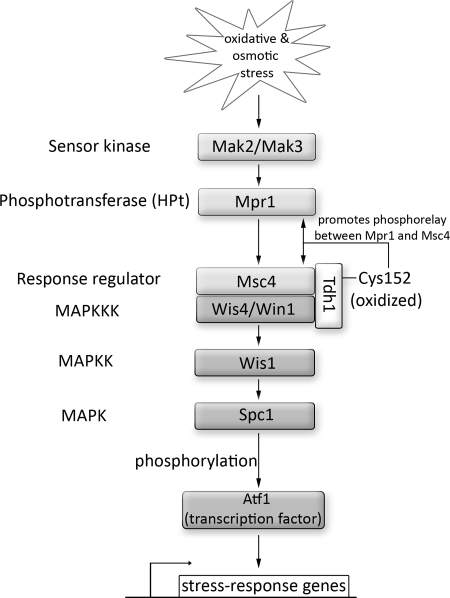
References
-
- Abate C. Patel L. Rauscher FJ., 3rd Curran T. Redox regulation of fos and jun DNA-binding activity in vitro. Science. 1990;249:1157–1161. - PubMed
-
- Allen RG. Tresini M. Oxidative stress and gene regulation. Free Radic Biol Med. 2000;28:463–499. - PubMed
-
- Alonso A. Sasin J. Bottini N. Friedberg I. Friedberg I. Osterman A. Godzik A. Hunter T. Dixon J. Mustelin T. Protein tyrosine phosphatases in the human genome. Cell. 2004;117:699–711. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
