

Spastin couples microtubule severing to membrane traffic in completion of cytokinesis and secretion
- PMID: 19000169
- PMCID: PMC2709849
- DOI: 10.1111/j.1600-0854.2008.00847.x
Spastin couples microtubule severing to membrane traffic in completion of cytokinesis and secretion
Abstract
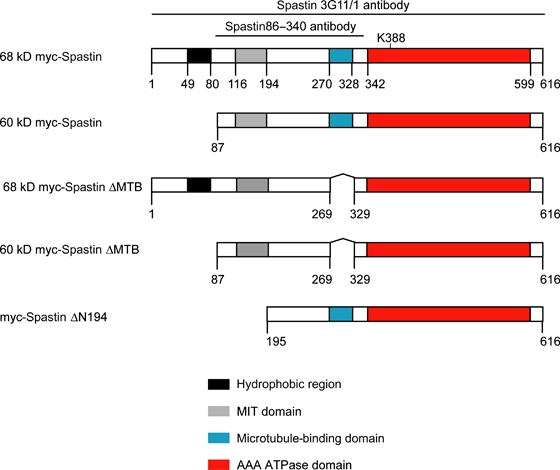
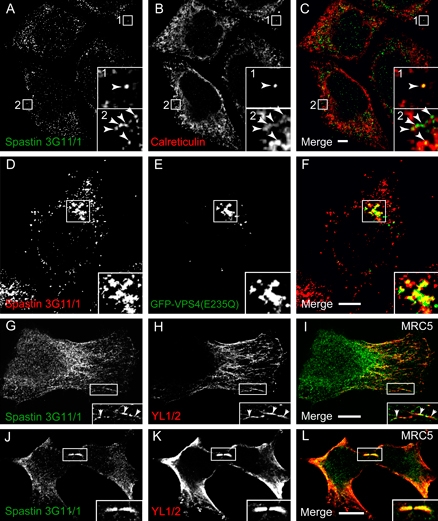
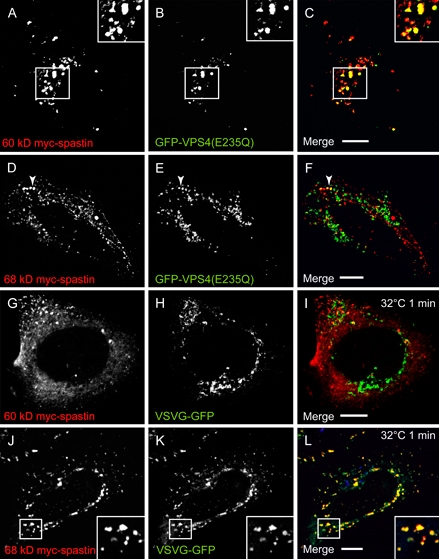
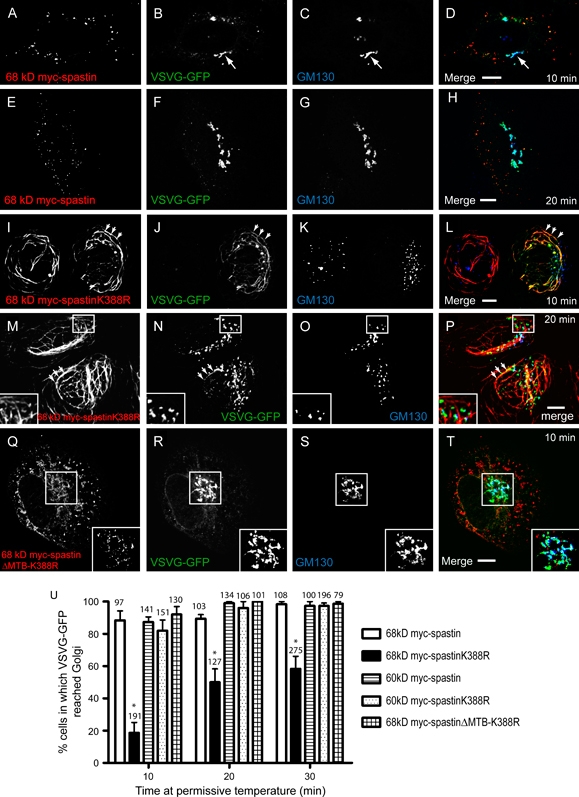
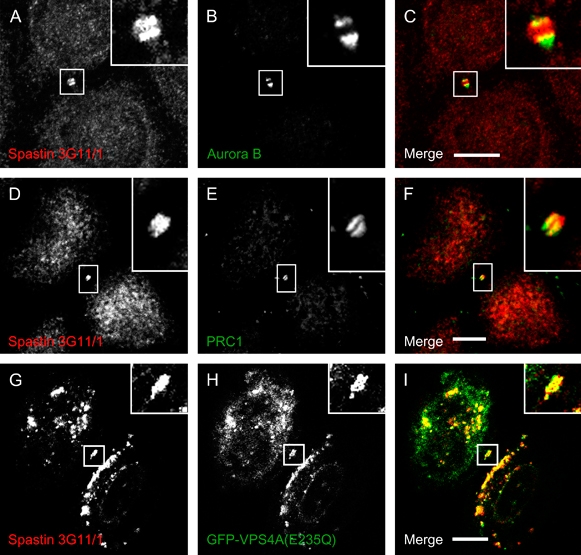
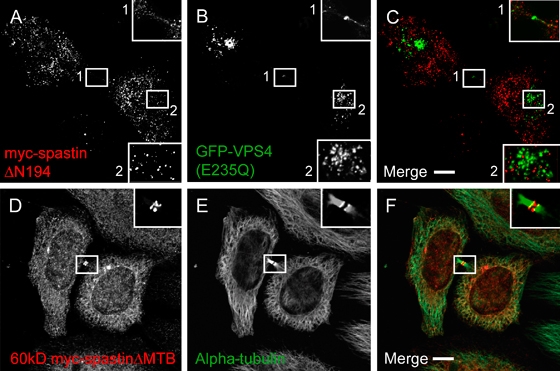
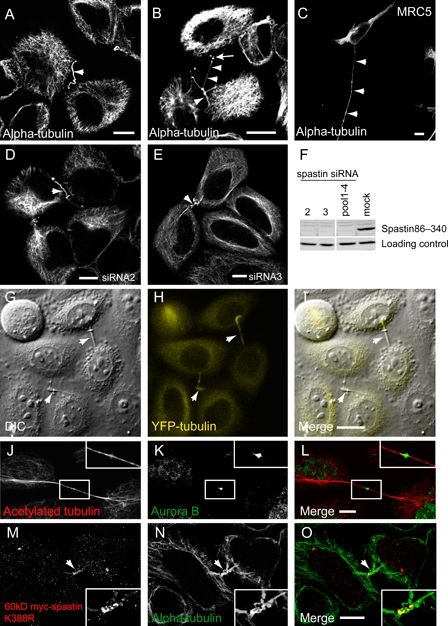
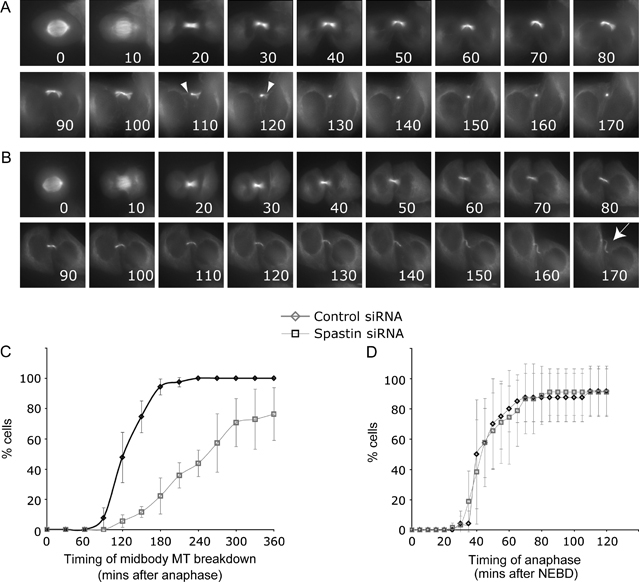
Mutations in the gene encoding the microtubule (MT)-severing protein spastin are the most common cause of hereditary spastic paraplegia, a genetic condition in which axons of the corticospinal tracts degenerate. We show that not only does endogenous spastin colocalize with MTs, but that it is also located on the early secretory pathway, can be recruited to endosomes and is present in the cytokinetic midbody. Spastin has two main isoforms, a 68 kD full-length isoform and a 60 kD short form. These two isoforms preferentially localize to different membrane traffic pathways with 68 kD spastin being principally located at the early secretory pathway, where it regulates endoplasmic reticulum-to-Golgi traffic. Sixty kiloDalton spastin is the major form recruited to endosomes and is also present in the midbody, where its localization requires the endosomal sorting complex required for transport-III-interacting MIT domain. Loss of midbody MTs accompanies the abscission stage of cytokinesis. In cells lacking spastin, a MT disruption event that normally accompanies abscission does not occur and abscission fails. We suggest that this event represents spastin-mediated MT severing. Our results support a model in which membrane traffic and MT regulation are coupled through spastin. This model is relevant in the axon, where there also is co-ordinated MT regulation and membrane traffic.
Figures
References
-
- Reid E. The hereditary spastic paraplegias. J Neurol. 1999;246:995–1003. - PubMed
-
- Fink JK. Hereditary spastic paraplegia. Neurol Clin. 2002;20:711–726. - PubMed
-
- Hazan J, Fonknechten N, Mavel D, Paternotte C, Samson D, Artiguenave F, Davoine CS, Cruaud C, Durr A, Wincker P, Brottier P, Cattolico L, Barbe V, Burgunder JM, Prud’homme JF, et al. Spastin, a new AAA protein, is altered in the most frequent form of autosomal dominant spastic paraplegia. Nat Genet. 1999;23:296–303. - PubMed
-
- Fonknechten N, Mavel D, Byrne P, Davoine CS, Cruaud C, Bonsch D, Samson D, Coutinho P, Hutchinson M, McMonagle P, Burgunder JM, Tartaglione A, Heinzlef O, Feki I, Deufel T, et al. Spectrum of SPG4 mutations in autosomal dominant spastic paraplegia. Hum Mol Genet. 2000;9:637–644. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
