

Checkpoint-dependent regulation of origin firing and replication fork movement in response to DNA damage in fission yeast
- PMID: 19001087
- PMCID: PMC2612511
- DOI: 10.1128/MCB.01319-08
Checkpoint-dependent regulation of origin firing and replication fork movement in response to DNA damage in fission yeast
Abstract
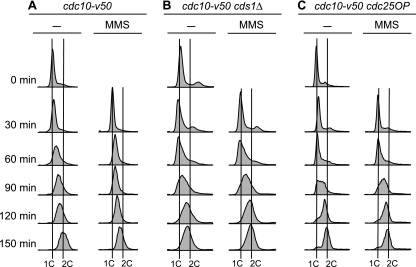
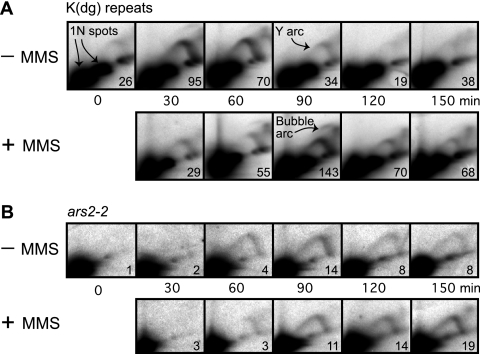
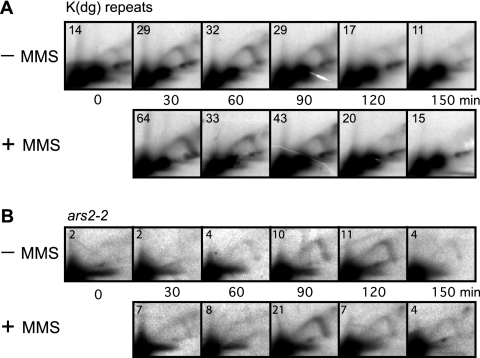
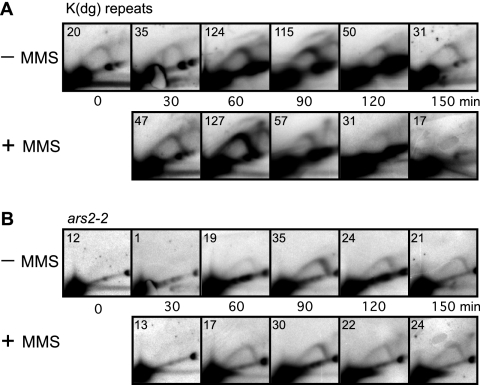
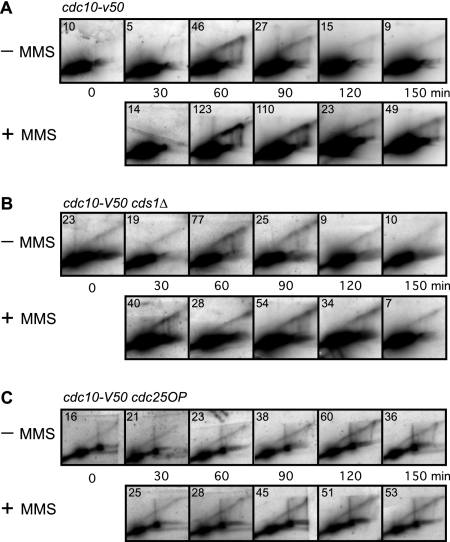
To elucidate the checkpoint mechanism responsible for slowing passage through S phase when fission yeast cells are treated with the DNA-damaging agent methyl methanesulfonate (MMS), we carried out two-dimensional gel analyses of replication intermediates in cells synchronized by cdc10 block (in G(1)) followed by release into synchronous S phase. The results indicated that under these conditions early-firing centromeric origins were partially delayed but late-firing telomeric origins were not delayed. Replication intermediates persisted in MMS-treated cells, suggesting that replication fork movement was inhibited. These effects were dependent on the Cds1 checkpoint kinase and were abolished in cells overexpressing the Cdc25 phosphatase, suggesting a role for the Cdc2 cyclin-dependent kinase. We conclude that both partial inhibition of the firing of a subset of origins and inhibition of replication fork movement contribute to the slowing of S phase in MMS-treated fission yeast cells.
Figures
References
-
- Alcasabas, A. A., A. J. Osborn, J. Bacchant, F. Hu, P. J. H. Werler, K. Bousset, K. Furuya, J. F. X. Diffley, A. M. Carr, and S. J. Elledge. 2001. Mrc1 transduces signals of DNA replication stress to activate Rad53. Nat. Cell Biol. 3958-965. - PubMed
-
- Andreassen, P. R., G. P. H. Ho, and A. D. D'Andrea. 2006. DNA damage responses and their many interactions with the replication fork. Carcinogenesis 27883-892. - PubMed
-
- Boddy, M. N., and P. Russell. 2001. DNA replication checkpoint. Curr. Biol. 11R953-R956. - PubMed
-
- Brewer, B. J., and W. L. Fangman. 1987. The localization of replication origins on ARS plasmids in S. cerevisiae. Cell 51463-471. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous