

Nicotinamide riboside and nicotinic acid riboside salvage in fungi and mammals. Quantitative basis for Urh1 and purine nucleoside phosphorylase function in NAD+ metabolism
- PMID: 19001417
- PMCID: PMC2610512
- DOI: 10.1074/jbc.M807976200
Nicotinamide riboside and nicotinic acid riboside salvage in fungi and mammals. Quantitative basis for Urh1 and purine nucleoside phosphorylase function in NAD+ metabolism
Erratum in
- J Biol Chem. 2009 Mar 20;284(12):8208
Abstract
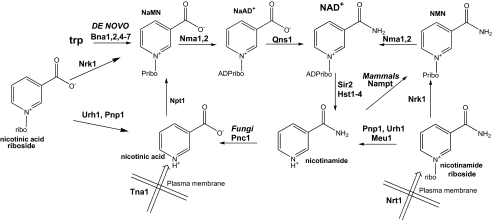
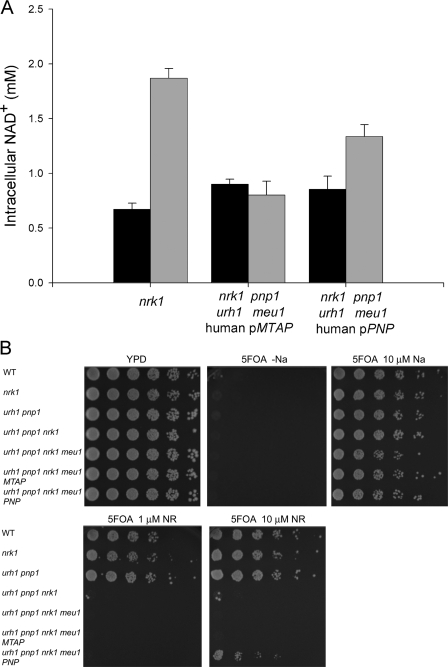
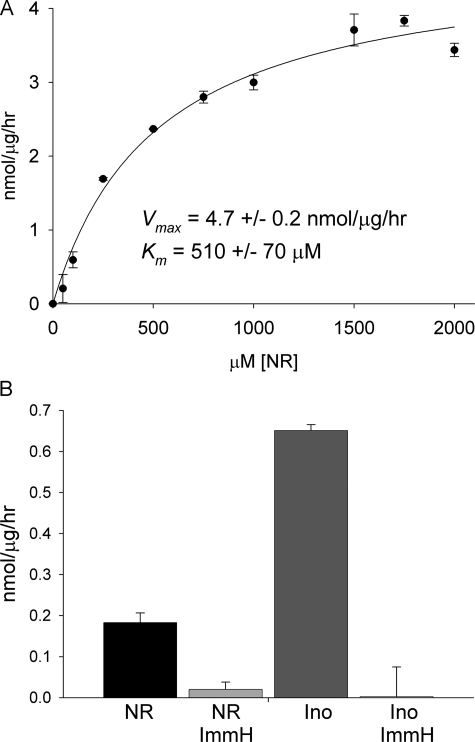
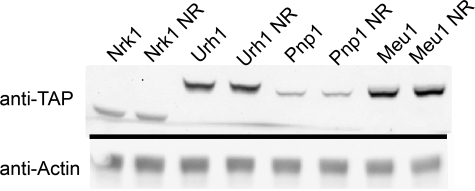
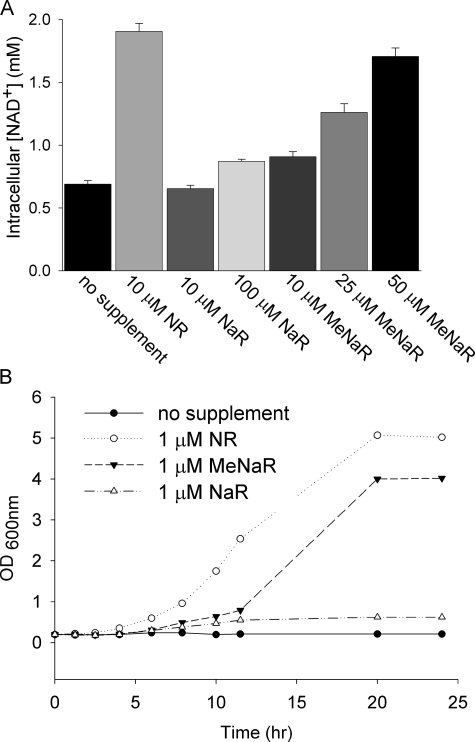
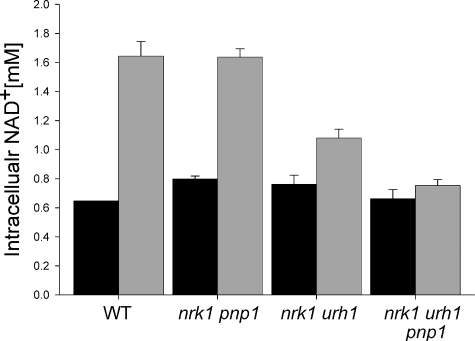
NAD+ is a co-enzyme for hydride transfer enzymes and an essential substrate of ADP-ribose transfer enzymes and sirtuins, the type III protein lysine deacetylases related to yeast Sir2. Supplementation of yeast cells with nicotinamide riboside extends replicative lifespan and increases Sir2-dependent gene silencing by virtue of increasing net NAD+ synthesis. Nicotinamide riboside elevates NAD+ levels via the nicotinamide riboside kinase pathway and by a pathway initiated by splitting the nucleoside into a nicotinamide base followed by nicotinamide salvage. Genetic evidence has established that uridine hydrolase, purine nucleoside phosphorylase, and methylthioadenosine phosphorylase are required for Nrk-independent utilization of nicotinamide riboside in yeast. Here we show that mammalian purine nucleoside phosphorylase but not methylthioadenosine phosphorylase is responsible for mammalian nicotinamide riboside kinase-independent nicotinamide riboside utilization. We demonstrate that so-called uridine hydrolase is 100-fold more active as a nicotinamide riboside hydrolase than as a uridine hydrolase and that uridine hydrolase and mammalian purine nucleoside phosphorylase cleave nicotinic acid riboside, whereas the yeast phosphorylase has little activity on nicotinic acid riboside. Finally, we show that yeast nicotinic acid riboside utilization largely depends on uridine hydrolase and nicotinamide riboside kinase and that nicotinic acid riboside bioavailability is increased by ester modification.
Figures
References
-
- Belenky, P., Bogan, K. L., and Brenner, C. (2007) Trends Biochem. Sci. 32 12–19 - PubMed
-
- Tanny, J. C., Dowd, G. J., Huang, J., Hilz, H., and Moazed, D. (1999) Cell 99 735–745 - PubMed
-
- Sauve, A. A., Celic, I., Avalos, J., Deng, H., Boeke, J. D., and Schramm, V. L. (2001) Biochemistry 40 15456–15463 - PubMed
-
- Guarente, L. (2000) Genes Dev. 14 1021–1026 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
