

Two distinct regions of Mto1 are required for normal microtubule nucleation and efficient association with the gamma-tubulin complex in vivo
- PMID: 19001497
- PMCID: PMC2743986
- DOI: 10.1242/jcs.038414
Two distinct regions of Mto1 are required for normal microtubule nucleation and efficient association with the gamma-tubulin complex in vivo
Abstract
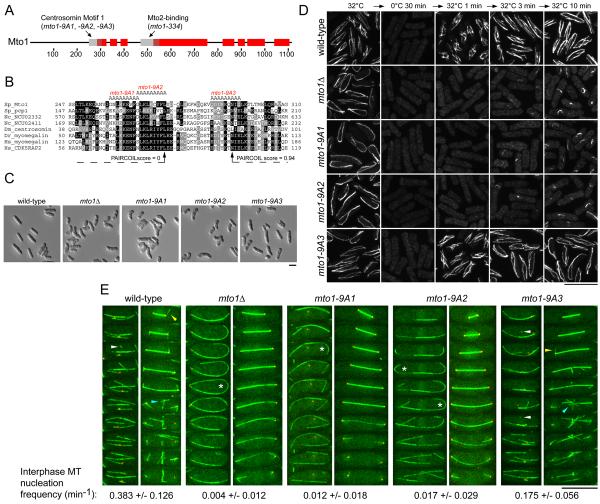
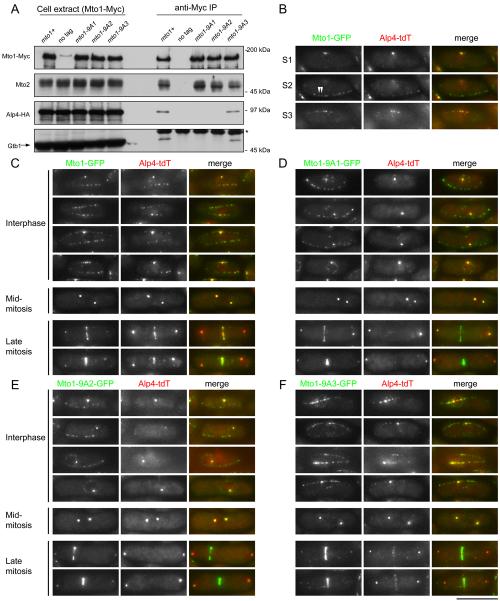
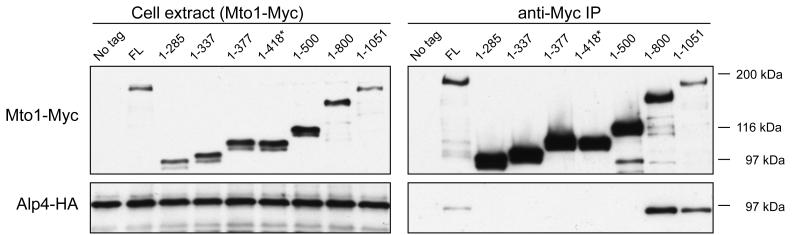
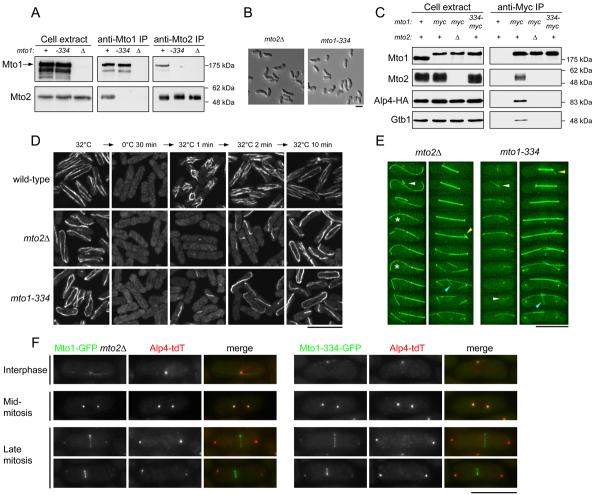
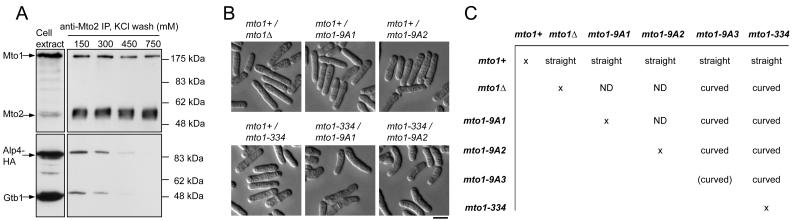
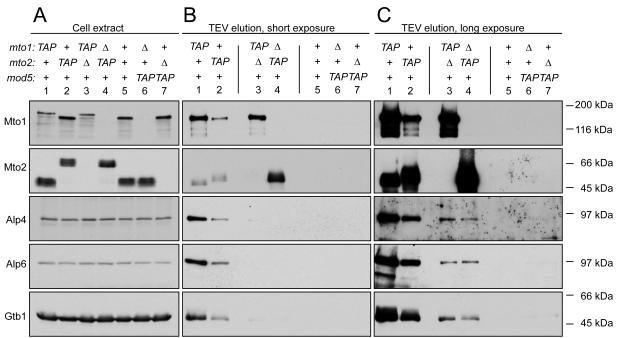
Cytoplasmic microtubule nucleation in the fission yeast Schizosaccharomyces pombe involves the interacting proteins Mto1 and Mto2, which are thought to recruit the gamma-tubulin complex (gamma-TuC) to prospective microtubule organizing centres. Mto1 contains a short amino-terminal region (CM1) that is conserved in higher eukaryotic proteins implicated in microtubule organization, centrosome function and/or brain development. Here we show that mutations in the Mto1 CM1 region generate mutant proteins that are functionally null for cytoplasmic microtubule nucleation and interaction with the gamma-TuC (phenocopying mto1Delta), even though the Mto1-mutant proteins localize normally in cells and can bind Mto2. Interestingly, the CM1 region is not sufficient for efficient interaction with the gamma-TuC. Mutation within a different region of Mto1, outside CM1, abrogates Mto2 binding and also impairs cytoplasmic microtubule nucleation and Mto1 association with the gamma-TuC. However, this mutation allows limited microtubule nucleation in vivo, phenocopying mto2Delta rather than mto1Delta. Further experiments suggest that Mto1 and Mto2 form a complex (Mto1/2 complex) independent of the gamma-TuC and that Mto1 and Mto2 can each associate with the gamma-TuC in the absence of the other, albeit extremely weakly compared to when both Mto1 and Mto2 are present. We propose that Mto2 acts cooperatively with Mto1 to promote association of the Mto1/2 complex with the gamma-TuC.
Figures
Similar articles
-
Activation of the γ-tubulin complex by the Mto1/2 complex.Curr Biol. 2014 Apr 14;24(8):896-903. doi: 10.1016/j.cub.2014.03.006. Epub 2014 Apr 3. Curr Biol. 2014. PMID: 24704079 Free PMC article.
-
Reconstitution of Microtubule Nucleation In Vitro Reveals Novel Roles for Mzt1.Curr Biol. 2019 Jul 8;29(13):2199-2207.e10. doi: 10.1016/j.cub.2019.05.058. Curr Biol. 2019. PMID: 31287970 Free PMC article.
-
Microtubule nucleation promoters Mto1 and Mto2 regulate cytokinesis in fission yeast.Mol Biol Cell. 2020 Aug 1;31(17):1846-1856. doi: 10.1091/mbc.E19-12-0686. Epub 2020 Jun 10. Mol Biol Cell. 2020. PMID: 32520628 Free PMC article.
-
Targeting of γ-tubulin complexes to microtubule organizing centers: conservation and divergence.Trends Cell Biol. 2015 May;25(5):296-307. doi: 10.1016/j.tcb.2014.12.002. Epub 2014 Dec 24. Trends Cell Biol. 2015. PMID: 25544667 Review.
-
Regulation of microtubule nucleation mediated by γ-tubulin complexes.Protoplasma. 2017 May;254(3):1187-1199. doi: 10.1007/s00709-016-1070-z. Epub 2017 Jan 10. Protoplasma. 2017. PMID: 28074286 Review.
Cited by
-
Cdk5rap2 exposes the centrosomal root of microcephaly syndromes.Trends Cell Biol. 2011 Aug;21(8):470-80. doi: 10.1016/j.tcb.2011.04.007. Epub 2011 May 31. Trends Cell Biol. 2011. PMID: 21632253 Free PMC article. Review.
-
Klp2-mediated Rsp1-Mto1 colocalization inhibits microtubule-dependent microtubule assembly in fission yeast.Sci Adv. 2025 Jan 3;11(1):eadq0670. doi: 10.1126/sciadv.adq0670. Epub 2025 Jan 3. Sci Adv. 2025. PMID: 39752482 Free PMC article.
-
Nuclear Mechanics in the Fission Yeast.Cells. 2019 Oct 20;8(10):1285. doi: 10.3390/cells8101285. Cells. 2019. PMID: 31635174 Free PMC article. Review.
-
Fission yeast Mto1 regulates diversity of cytoplasmic microtubule organizing centers.Curr Biol. 2010 Nov 9;20(21):1959-65. doi: 10.1016/j.cub.2010.10.006. Epub 2010 Oct 21. Curr Biol. 2010. PMID: 20970338 Free PMC article.
-
Fission yeast MOZART1/Mzt1 is an essential γ-tubulin complex component required for complex recruitment to the microtubule organizing center, but not its assembly.Mol Biol Cell. 2013 Sep;24(18):2894-906. doi: 10.1091/mbc.E13-05-0235. Epub 2013 Jul 24. Mol Biol Cell. 2013. PMID: 23885124 Free PMC article.
References
-
- Bahler J, Wu JQ, Longtine MS, Shah NG, McKenzie A, 3rd, Steever AB, Wach A, Philippsen P, Pringle JR. Heterologous modules for efficient and versatile PCR-based gene targeting in Schizosaccharomyces pombe. Yeast. 1998;14:943–51. - PubMed
-
- Bond J, Roberts E, Springell K, Lizarraga SB, Scott S, Higgins J, Hampshire DJ, Morrison EE, Leal GF, Silva EO, et al. A centrosomal mechanism involving CDK5RAP2 and CENPJ controls brain size. Nat. Genet. 2005;37:353–5. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
