

Foamy macrophages from tuberculous patients' granulomas constitute a nutrient-rich reservoir for M. tuberculosis persistence
- PMID: 19002241
- PMCID: PMC2575403
- DOI: 10.1371/journal.ppat.1000204
Foamy macrophages from tuberculous patients' granulomas constitute a nutrient-rich reservoir for M. tuberculosis persistence
Abstract
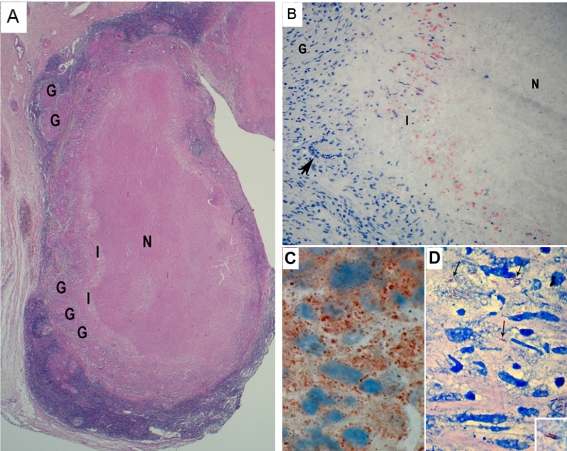
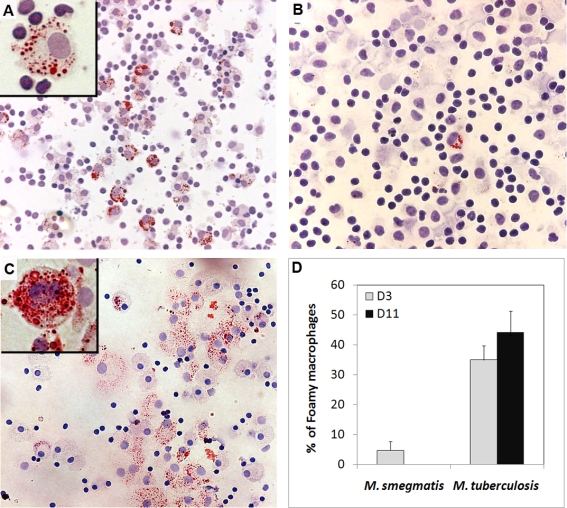
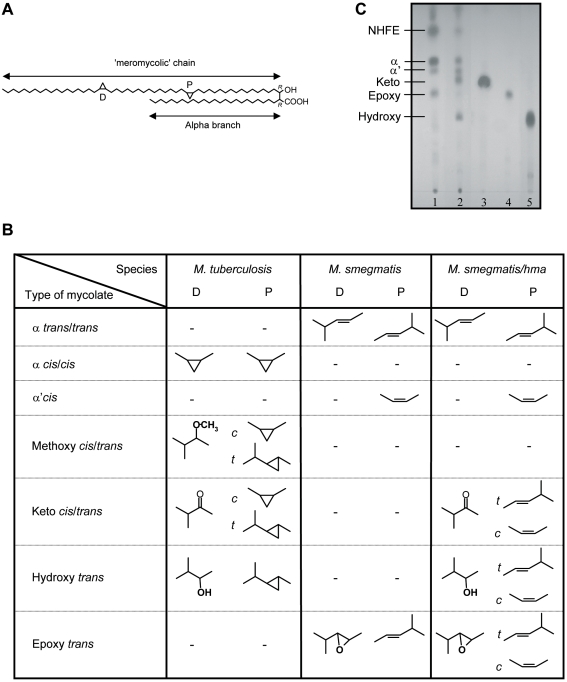
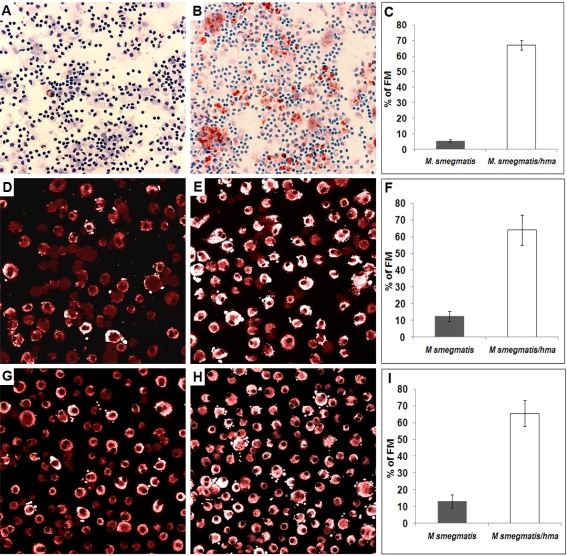
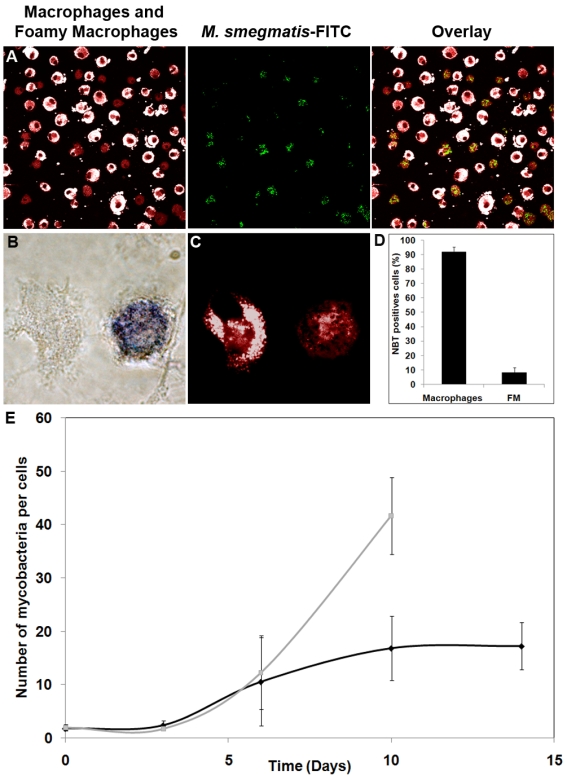
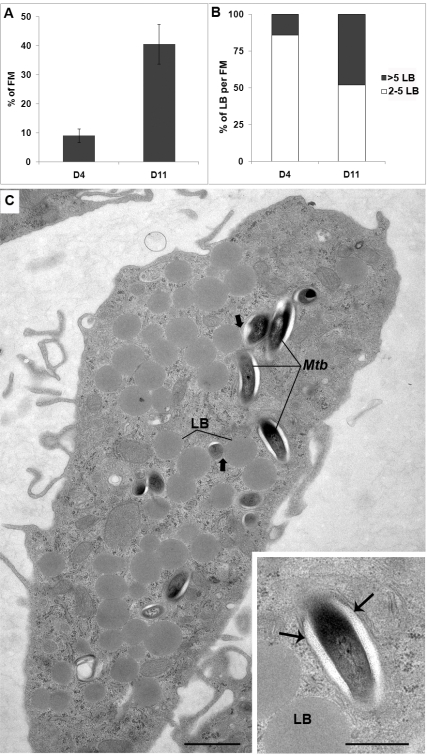
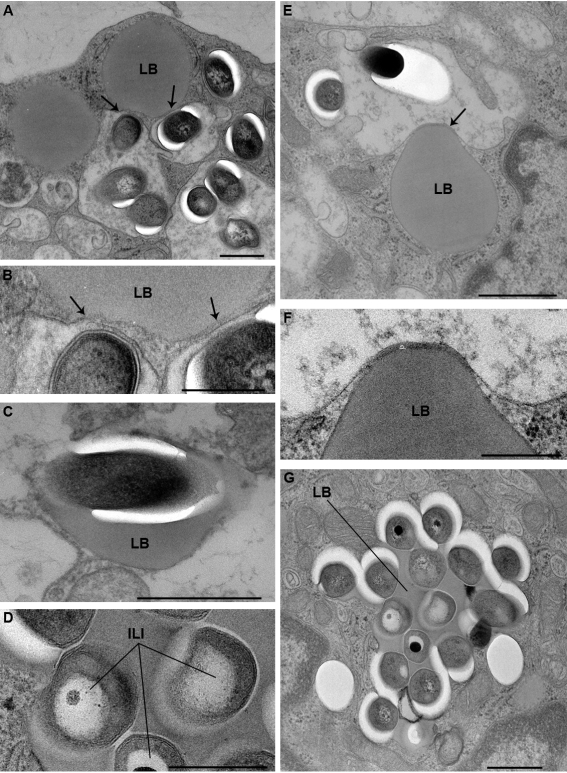
Tuberculosis (TB) is characterized by a tight interplay between Mycobacterium tuberculosis and host cells within granulomas. These cellular aggregates restrict bacterial spreading, but do not kill all the bacilli, which can persist for years. In-depth investigation of M. tuberculosis interactions with granuloma-specific cell populations are needed to gain insight into mycobacterial persistence, and to better understand the physiopathology of the disease. We have analyzed the formation of foamy macrophages (FMs), a granuloma-specific cell population characterized by its high lipid content, and studied their interaction with the tubercle bacillus. Within our in vitro human granuloma model, M. tuberculosis long chain fatty acids, namely oxygenated mycolic acids (MA), triggered the differentiation of human monocyte-derived macrophages into FMs. In these cells, mycobacteria no longer replicated and switched to a dormant non-replicative state. Electron microscopy observation of M. tuberculosis-infected FMs showed that the mycobacteria-containing phagosomes migrate towards host cell lipid bodies (LB), a process which culminates with the engulfment of the bacillus into the lipid droplets and with the accumulation of lipids within the microbe. Altogether, our results suggest that oxygenated mycolic acids from M. tuberculosis play a crucial role in the differentiation of macrophages into FMs. These cells might constitute a reservoir used by the tubercle bacillus for long-term persistence within its human host, and could provide a relevant model for the screening of new antimicrobials against non-replicating persistent mycobacteria.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- WHO. Global Tuberculosis Control, WHO Report. 2006. http://www.who.int/mediacentre/factsheets/fs104/en/index.html.
-
- Glickman MS, Jacobs WR., Jr Microbial pathogenesis of Mycobacterium tuberculosis: dawn of a discipline. Cell. 2001;104:477–485. - PubMed
-
- Tufariello JM, Chan J, Flynn JL. Latent tuberculosis: mechanisms of host and bacillus that contribute to persistent infection. Lancet Infect Dis. 2003;3:578–590. - PubMed
-
- Cardona PJ, Llatjos R, Gordillo S, Diaz J, Ojanguren I, et al. Evolution of granulomas in lungs of mice infected aerogenially with Mycobacterium tuberculosis. Scand J Immunol. 2000;52:156–163. - PubMed
-
- Ridley DS, Ridley MJ. Rationale for the histological spectrum of tuberculosis: a basis for classification. Pathology. 1987;19:186–192. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
