

Bacillus anthracis peptidoglycan stimulates an inflammatory response in monocytes through the p38 mitogen-activated protein kinase pathway
- PMID: 19002259
- PMCID: PMC2577892
- DOI: 10.1371/journal.pone.0003706
Bacillus anthracis peptidoglycan stimulates an inflammatory response in monocytes through the p38 mitogen-activated protein kinase pathway
Abstract
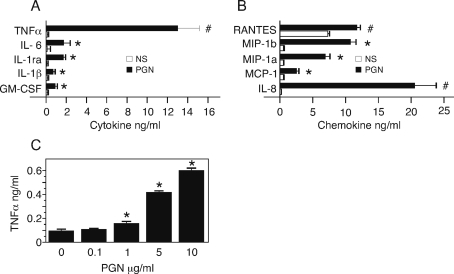
We hypothesized that the peptidoglycan component of B. anthracis may play a critical role in morbidity and mortality associated with inhalation anthrax. To explore this issue, we purified the peptidoglycan component of the bacterial cell wall and studied the response of human peripheral blood cells. The purified B. anthracis peptidoglycan was free of non-covalently bound protein but contained a complex set of amino acids probably arising from the stem peptide. The peptidoglycan contained a polysaccharide that was removed by mild acid treatment, and the biological activity remained with the peptidoglycan and not the polysaccharide. The biological activity of the peptidoglycan was sensitive to lysozyme but not other hydrolytic enzymes, showing that the activity resides in the peptidoglycan component and not bacterial DNA, RNA or protein. B. anthracis peptidoglycan stimulated monocytes to produce primarily TNFalpha; neutrophils and lymphocytes did not respond. Peptidoglycan stimulated monocyte p38 mitogen-activated protein kinase and p38 activity was required for TNFalpha production by the cells. We conclude that peptidoglycan in B. anthracis is biologically active, that it stimulates a proinflammatory response in monocytes, and uses the p38 kinase signal transduction pathway to do so. Given the high bacterial burden in pulmonary anthrax, these findings suggest that the inflammatory events associated with peptidoglycan may play an important role in anthrax pathogenesis.
Conflict of interest statement
Figures






Similar articles
-
Bacillus anthracis' lethal toxin induces broad transcriptional responses in human peripheral monocytes.BMC Immunol. 2012 Jul 2;13:33. doi: 10.1186/1471-2172-13-33. BMC Immunol. 2012. PMID: 22747600 Free PMC article.
-
Inflammatory cytokine response to Bacillus anthracis peptidoglycan requires phagocytosis and lysosomal trafficking.Infect Immun. 2010 Jun;78(6):2418-28. doi: 10.1128/IAI.00170-10. Epub 2010 Mar 22. Infect Immun. 2010. PMID: 20308305 Free PMC article.
-
The function of mitogen-activated protein kinase phosphatase-1 in peptidoglycan-stimulated macrophages.J Biol Chem. 2004 Dec 24;279(52):54023-31. doi: 10.1074/jbc.M408444200. Epub 2004 Oct 13. J Biol Chem. 2004. PMID: 15485842
-
A review of the interaction of Bacillus anthracis with cells of the innate immune response.Berl Munch Tierarztl Wochenschr. 2006 May-Jun;119(5-6):216-21. Berl Munch Tierarztl Wochenschr. 2006. PMID: 16729468 Review.
-
Anthrax toxin.Crit Rev Microbiol. 2001;27(3):167-200. doi: 10.1080/20014091096738. Crit Rev Microbiol. 2001. PMID: 11596878 Review.
Cited by
-
Cellular and physiological effects of anthrax exotoxin and its relevance to disease.Front Cell Infect Microbiol. 2012 Jun 1;2:76. doi: 10.3389/fcimb.2012.00076. eCollection 2012. Front Cell Infect Microbiol. 2012. PMID: 22919667 Free PMC article. Review.
-
Assessment of immunotoxicity using precision-cut tissue slices.Xenobiotica. 2013 Jan;43(1):84-97. doi: 10.3109/00498254.2012.731543. Epub 2012 Nov 16. Xenobiotica. 2013. PMID: 23199366 Free PMC article. Review.
-
Serum Amyloid P and IgG Exhibit Differential Capabilities in the Activation of the Innate Immune System in Response to Bacillus anthracis Peptidoglycan.Infect Immun. 2018 Apr 23;86(5):e00076-18. doi: 10.1128/IAI.00076-18. Print 2018 May. Infect Immun. 2018. PMID: 29531132 Free PMC article.
-
Toxin inhibition of antimicrobial factors induced by Bacillus anthracis peptidoglycan in human blood.Infect Immun. 2013 Oct;81(10):3693-702. doi: 10.1128/IAI.00709-13. Epub 2013 Jul 22. Infect Immun. 2013. PMID: 23876807 Free PMC article.
-
C3 Opsonization of Anthrax Bacterium and Peptidoglycan Supports Recognition and Activation of Neutrophils.Microorganisms. 2020 Jul 13;8(7):1039. doi: 10.3390/microorganisms8071039. Microorganisms. 2020. PMID: 32668703 Free PMC article.
References
-
- Dixon TC, Meselson M, Guillemin J, Hanna PC. Anthrax. N Engl J Med. 1999;341:815–826. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
