

Mechanisms of minocycline-induced suppression of simian immunodeficiency virus encephalitis: inhibition of apoptosis signal-regulating kinase 1
- PMID: 19003592
- PMCID: PMC2627770
- DOI: 10.1080/13550280802199898
Mechanisms of minocycline-induced suppression of simian immunodeficiency virus encephalitis: inhibition of apoptosis signal-regulating kinase 1
Abstract
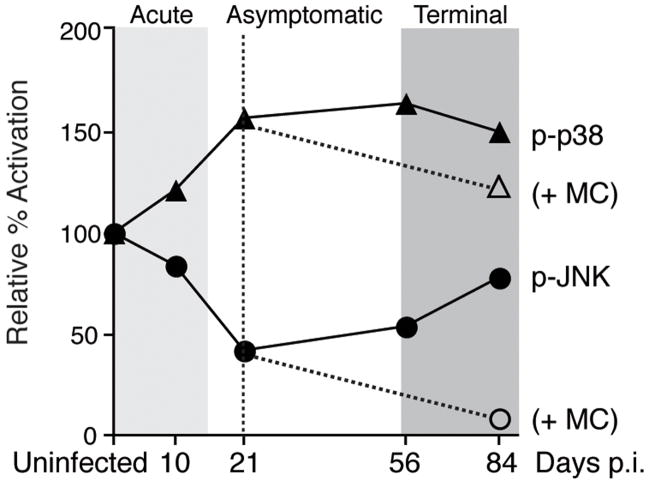
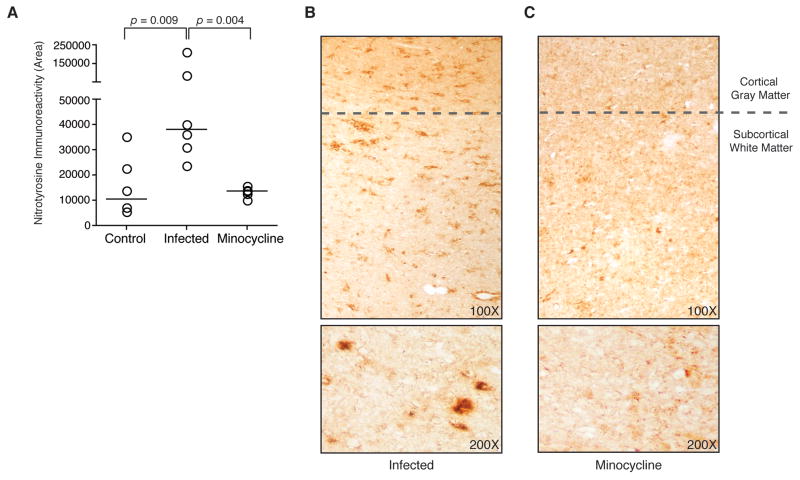
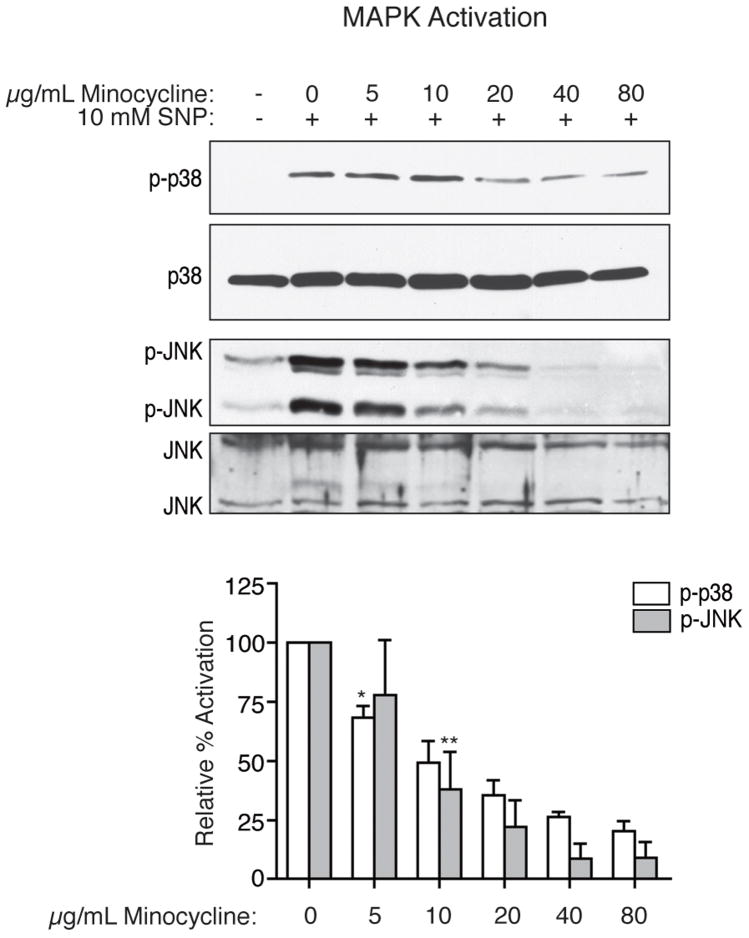
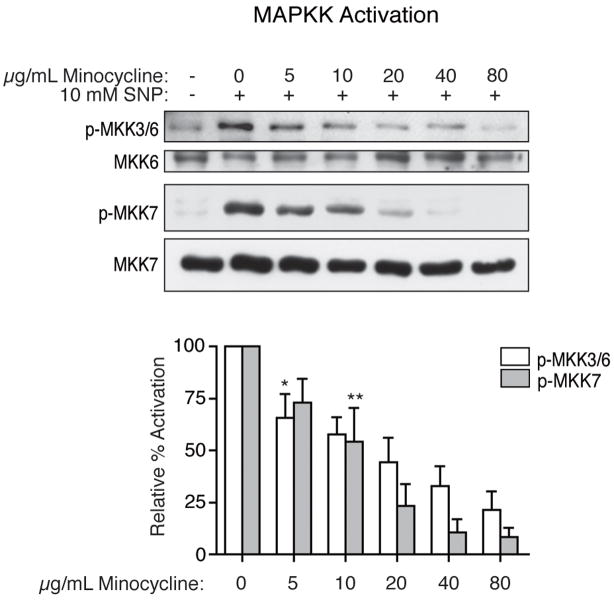
Human immunodeficiency virus (HIV) infection of the central nervous system (CNS) can lead to cognitive dysfunction, even in individuals treated with highly active antiretroviral therapy. Using an established simian immunodeficiency virus (SIV)/macaque model of HIV CNS disease, we previously reported that infection shifts the balance of activation of mitogen-activated protein kinase (MAPK) signaling pathways in the brain, resulting in increased activation of the neurodegenerative MAPKs p38 and JNK. Minocycline treatment of SIV-infected macaques reduced the incidence and severity of SIV encephalitis in this model, and suppressed the activation of p38 in the brain. The purpose of this study was to further examine the effects of minocycline on neurodegenerative MAPK signaling. We first demonstrated that minocycline also decreases JNK activation in the brain and levels of the inflammatory mediator nitric oxide (NO). We next used NO to activate these MAPK pathways in vitro, and demonstrated that minocycline suppresses p38 and c-Jun N-terminal kinase (JNK) activation by reducing intracellular levels, and hence, activation of apoptosis signal-regulating kinase 1 (ASK1), a MAPK kinase capable of selectively activating both pathways. We then demonstrated that ASK1 activation in the brain during SIV infection is suppressed by minocycline. By suppressing p38 and JNK activation pathways, which are important for the production of and responses to inflammatory mediators, minocycline may interrupt the vicious cycle of inflammation that both results from, and promotes, virus replication in SIV and HIV CNS disease.
Figures



Similar articles
-
Neuroprotective and anti-human immunodeficiency virus activity of minocycline.JAMA. 2005 Apr 27;293(16):2003-11. doi: 10.1001/jama.293.16.2003. JAMA. 2005. PMID: 15855434
-
Dysregulation of mitogen-activated protein kinase signaling pathways in simian immunodeficiency virus encephalitis.Am J Pathol. 2004 Feb;164(2):355-62. doi: 10.1016/S0002-9440(10)63125-2. Am J Pathol. 2004. PMID: 14742241 Free PMC article.
-
Brain Macrophages in Simian Immunodeficiency Virus-Infected, Antiretroviral-Suppressed Macaques: a Functional Latent Reservoir.mBio. 2017 Aug 15;8(4):e01186-17. doi: 10.1128/mBio.01186-17. mBio. 2017. PMID: 28811349 Free PMC article.
-
An SIV macaque model of SIV and HAND: the need for adjunctive therapies in HIV that target activated monocytes and macrophages.J Neurovirol. 2018 Apr;24(2):213-219. doi: 10.1007/s13365-018-0616-6. Epub 2018 Feb 12. J Neurovirol. 2018. PMID: 29435829 Review.
-
Activation mechanisms of ASK1 in response to various stresses and its significance in intracellular signaling.Adv Biol Regul. 2013 Jan;53(1):135-44. doi: 10.1016/j.jbior.2012.09.006. Epub 2012 Sep 13. Adv Biol Regul. 2013. PMID: 23031789 Review.
Cited by
-
Use of minocycline in viral infections.Indian J Med Res. 2011 May;133(5):467-70. Indian J Med Res. 2011. PMID: 21623029 Free PMC article. Review.
-
HIV-related neurocognitive impairment in the HAART era.Curr HIV/AIDS Rep. 2009 Aug;6(3):146-52. doi: 10.1007/s11904-009-0020-1. Curr HIV/AIDS Rep. 2009. PMID: 19589300 Review.
-
Unraveling the pathogenesis of HIV peripheral neuropathy: insights from a simian immunodeficiency virus macaque model.ILAR J. 2014;54(3):296-303. doi: 10.1093/ilar/ilt047. ILAR J. 2014. PMID: 24615443 Free PMC article. Review.
-
Minocycline attenuates HIV-1 infection and suppresses chronic immune activation in humanized NOD/LtsZ-scidIL-2Rγ(null) mice.Immunology. 2014 Aug;142(4):562-72. doi: 10.1111/imm.12246. Immunology. 2014. PMID: 24409837 Free PMC article.
-
Neurovirulent and non-neurovirulent strains of HIV-1 and their Tat proteins induce differential cytokine-chemokine profiles.NeuroImmune Pharm Ther. 2025 Feb 10;4(1):39-47. doi: 10.1515/nipt-2024-0020. eCollection 2025 Mar. NeuroImmune Pharm Ther. 2025. PMID: 40313367 Free PMC article.
References
-
- Akterin S, Cowburn RF, Miranda-Vizuete A, Jimenez A, Bogdanovic N, Winblad B, Cedazo-Minguez A. Involvement of glutaredoxin-1 and thioredoxin-1 in beta-amyloid toxicity and Alzheimer’s disease. Cell Death Differ. 2006;13:1454–65. - PubMed
-
- Ances BM, Ellis RJ. Dementia and neurocognitive disorders due to HIV-1 infection. Semin Neurol. 2007;27:86–92. - PubMed
-
- Borderie D, Hernvann A, Hilliquin P, Lemarchal H, Kahan A, Ekindjian OG. Tetracyclines inhibit nitrosothiol production by cytokine-stimulated osteoarthritic synovial cells. Inflamm Res. 2001;50:409–14. - PubMed
-
- Borsello T, Forloni G. JNK signalling: a possible target to prevent neurodegeneration. Curr Pharm Des. 2007;13:1875–86. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
