

Differential degradation of PIN2 auxin efflux carrier by retromer-dependent vacuolar targeting
- PMID: 19004783
- PMCID: PMC2584678
- DOI: 10.1073/pnas.0808073105
Differential degradation of PIN2 auxin efflux carrier by retromer-dependent vacuolar targeting
Abstract
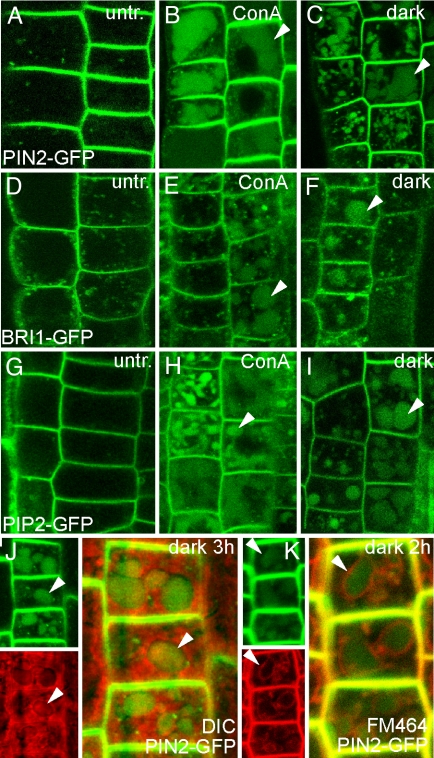
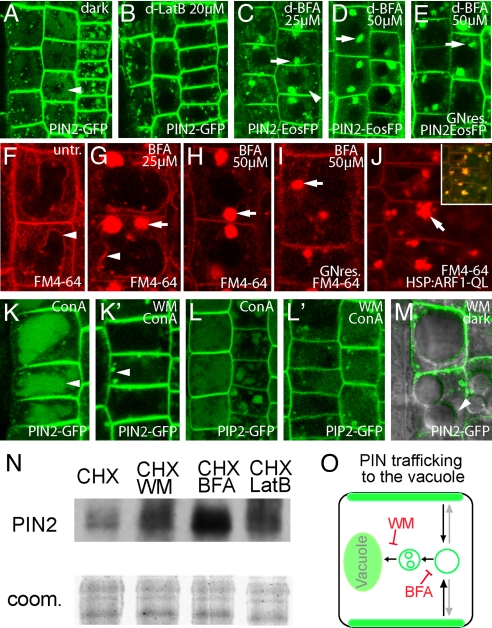
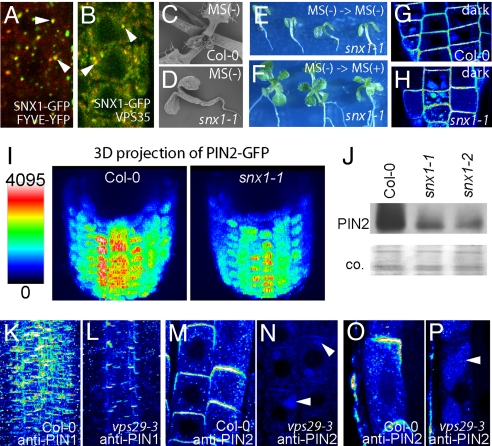
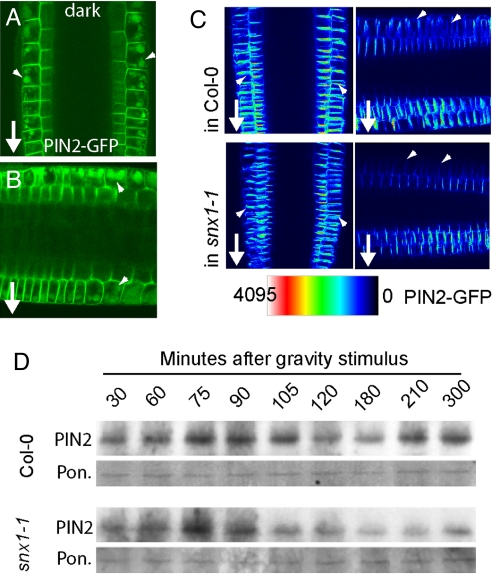
All eukaryotic cells present at the cell surface a specific set of plasma membrane proteins that modulate responses to internal and external cues and whose activity is also regulated by protein degradation. We characterized the lytic vacuole-dependent degradation of membrane proteins in Arabidopsis thaliana by means of in vivo visualization of vacuolar targeting combined with quantitative protein analysis. We show that the vacuolar targeting pathway is used by multiple cargos including PIN-FORMED (PIN) efflux carriers for the phytohormone auxin. In vivo visualization of PIN2 vacuolar targeting revealed its differential degradation in response to environmental signals, such as gravity. In contrast to polar PIN delivery to the basal plasma membrane, which depends on the vesicle trafficking regulator ARF-GEF GNOM, PIN sorting to the lytic vacuolar pathway requires additional brefeldin A-sensitive ARF-GEF activity. Furthermore, we identified putative retromer components SORTING NEXIN1 (SNX1) and VACUOLAR PROTEIN SORTING29 (VPS29) as important factors in this pathway and propose that the retromer complex acts to retrieve PIN proteins from a late/pre-vacuolar compartment back to the recycling pathways. Our data suggest that ARF GEF- and retromer-dependent processes regulate PIN sorting to the vacuole in an antagonistic manner and illustrate instrumentalization of this mechanism for fine-tuning the auxin fluxes during gravitropic response.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
PIN auxin efflux carrier polarity is regulated by PINOID kinase-mediated recruitment into GNOM-independent trafficking in Arabidopsis.Plant Cell. 2009 Dec;21(12):3839-49. doi: 10.1105/tpc.109.071639. Epub 2009 Dec 29. Plant Cell. 2009. PMID: 20040538 Free PMC article.
-
BEX1/ARF1A1C is required for BFA-sensitive recycling of PIN auxin transporters and auxin-mediated development in Arabidopsis.Plant Cell Physiol. 2014 Apr;55(4):737-49. doi: 10.1093/pcp/pct196. Epub 2013 Dec 24. Plant Cell Physiol. 2014. PMID: 24369434 Free PMC article.
-
SCF(TIR1/AFB)-auxin signalling regulates PIN vacuolar trafficking and auxin fluxes during root gravitropism.EMBO J. 2013 Jan 23;32(2):260-74. doi: 10.1038/emboj.2012.310. Epub 2012 Dec 4. EMBO J. 2013. PMID: 23211744 Free PMC article.
-
Subcellular trafficking of PIN auxin efflux carriers in auxin transport.Eur J Cell Biol. 2010 Feb-Mar;89(2-3):231-5. doi: 10.1016/j.ejcb.2009.11.003. Epub 2009 Nov 26. Eur J Cell Biol. 2010. PMID: 19944476 Review.
-
Role of the GNOM gene in Arabidopsis apical-basal patterning--From mutant phenotype to cellular mechanism of protein action.Eur J Cell Biol. 2010 Feb-Mar;89(2-3):138-44. doi: 10.1016/j.ejcb.2009.11.020. Epub 2009 Dec 29. Eur J Cell Biol. 2010. PMID: 20036441 Review.
Cited by
-
Molecular mechanisms of gravity perception and signal transduction in plants.Protoplasma. 2016 Jul;253(4):987-1004. doi: 10.1007/s00709-015-0859-5. Epub 2015 Jul 28. Protoplasma. 2016. PMID: 26215561 Review.
-
ECHIDNA-mediated post-Golgi trafficking of auxin carriers for differential cell elongation.Proc Natl Acad Sci U S A. 2013 Oct 1;110(40):16259-64. doi: 10.1073/pnas.1309057110. Epub 2013 Sep 16. Proc Natl Acad Sci U S A. 2013. PMID: 24043780 Free PMC article.
-
The plant-unique protein DRIF1 coordinates with sorting nexin 1 to regulate membrane protein homeostasis.Plant Cell. 2023 Nov 30;35(12):4217-4237. doi: 10.1093/plcell/koad227. Plant Cell. 2023. PMID: 37647529 Free PMC article.
-
WRKY23 is a component of the transcriptional network mediating auxin feedback on PIN polarity.PLoS Genet. 2018 Jan 29;14(1):e1007177. doi: 10.1371/journal.pgen.1007177. eCollection 2018 Jan. PLoS Genet. 2018. PMID: 29377885 Free PMC article.
-
VPS36-Mediated plasma membrane protein turnover is critical for Arabidopsis root gravitropism.Plant Signal Behav. 2017 Apr 3;12(4):e1307495. doi: 10.1080/15592324.2017.1307495. Plant Signal Behav. 2017. PMID: 28340324 Free PMC article.
References
-
- Petrášek J, et al. PIN proteins perform a rate-limiting function in cellular auxin efflux. Science. 2006;312:914–918. - PubMed
-
- Wišniewska J, et al. Polar PIN localization directs auxin flow in plants. Science. 2006;312:883. - PubMed
-
- Leyser O. Dynamic integration of auxin transport and signalling. Curr Biol. 2006;16:R424–R433. - PubMed
-
- Kleine-Vehn J, Friml J. Polar targeting and endocytic recycling in auxin-dependent plant development. Annu Rev Cell Dev Biol. 2008;24:447–473. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
