

Depurination within the intergenic region of Brome mosaic virus RNA3 inhibits viral replication in vitro and in vivo
- PMID: 19004869
- PMCID: PMC2602774
- DOI: 10.1093/nar/gkn896
Depurination within the intergenic region of Brome mosaic virus RNA3 inhibits viral replication in vitro and in vivo
Abstract
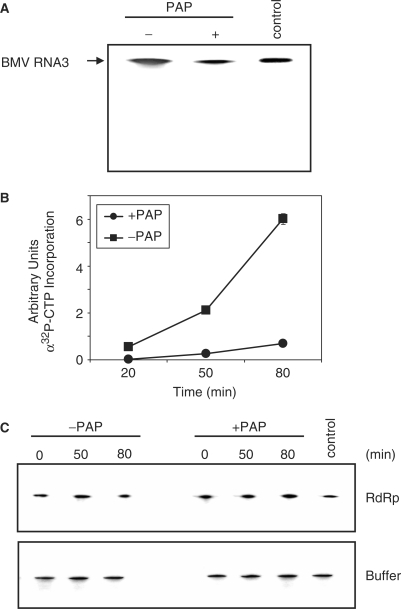
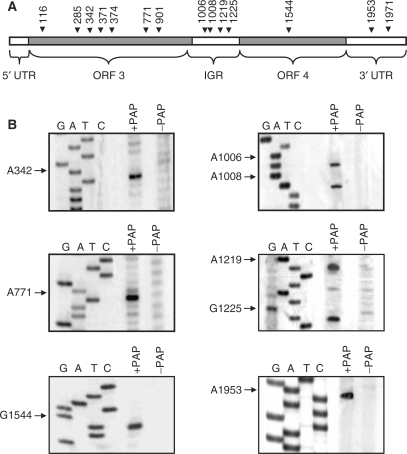
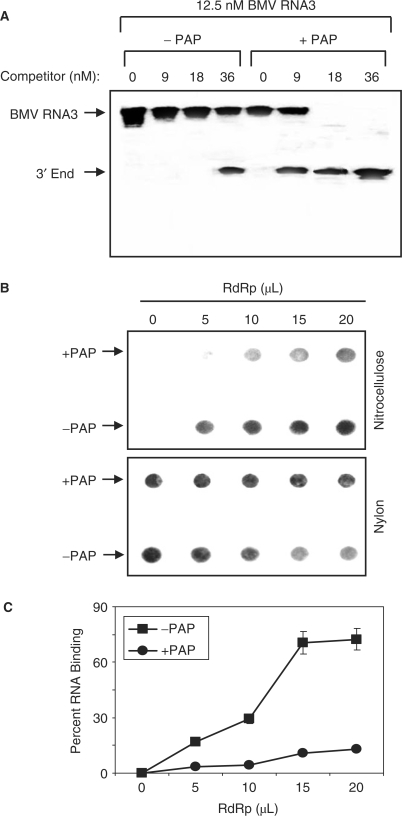
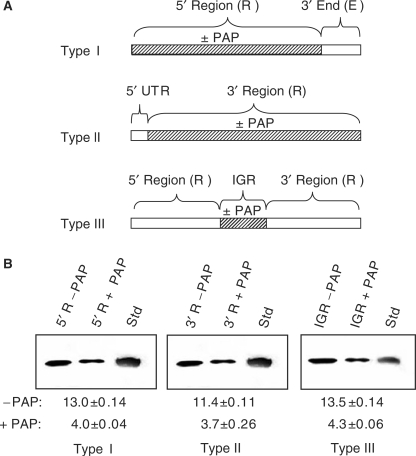
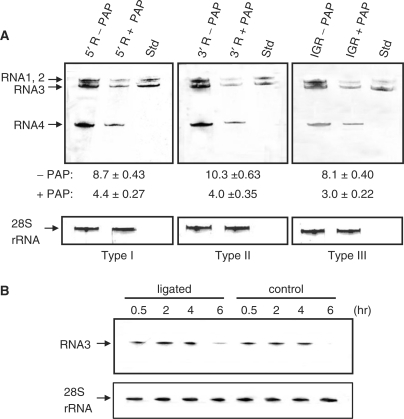
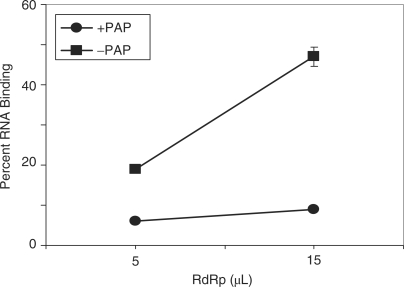
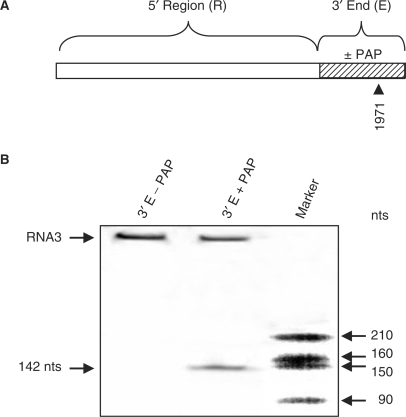
Pokeweed antiviral protein (PAP) is a glycosidase of plant origin that has been shown to depurinate some viral RNAs in vitro. We have demonstrated previously that treatment of Brome mosaic virus (BMV) RNAs with PAP inhibited their translation in a cell-free system and decreased their accumulation in barley protoplasts. In the current study, we map the depurination sites on BMV RNA3 and describe the mechanism by which replication of the viral RNA is inhibited by depurination. Specifically, we demonstrate that the viral replicase exhibited reduced affinity for depurinated positive-strand RNA3 compared with intact RNA3, resulting in less negative-strand product. This decrease was due to depurination within the intergenic region of RNA3, between ORF3 and 4, and distant from the 3' terminal core promoter required for initiation of negative-strand RNA synthesis. Depurination within the intergenic region alone inhibited the binding of the replicase to full-length RNA3, whereas depurination outside the intergenic region permitted the replicase to initiate negative-strand synthesis; however, elongation of the RNA product was stalled at the abasic nucleotide. These results support a role of the intergenic region in controlling negative-strand RNA synthesis and contribute new insight into the effect of depurination by PAP on BMV replication.
Figures
References
-
- Goldbach R, LeGall O, Wellink J. Alpha-like viruses in plants. Semin. Virol. 1991;2:19–25.
-
- Ahlquist P. Bromovirus RNA replication and transcription. Curr. Opin. Genet. Dev. 1992;2:71–76. - PubMed
-
- Kao CC, Sivakumaran K. Brome mosaic virus, good for an RNA virologist's basic needs. Mol. Plant Pathol. 2000;1:91–97. - PubMed
-
- Kong F, Sivakumaran K, Kao CC. The N-terminal half of the brome mosaic virus 1a protein has RNA capping associated activities: specificity for GTP and S-adenosylmethionine. Virology. 1999;259:200–210. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
