

A diversity of uncharacterized reverse transcriptases in bacteria
- PMID: 19004871
- PMCID: PMC2602772
- DOI: 10.1093/nar/gkn867
A diversity of uncharacterized reverse transcriptases in bacteria
Abstract
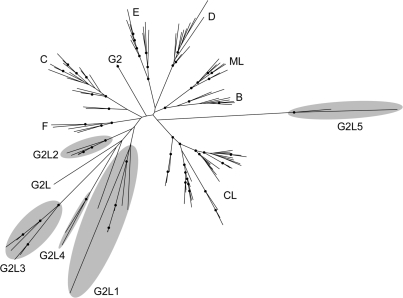
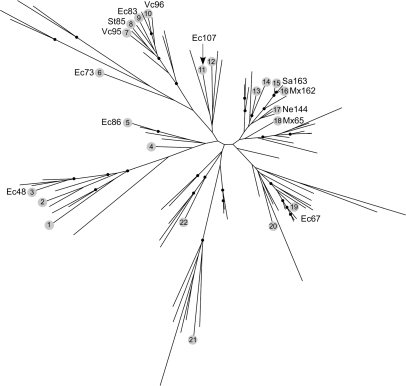
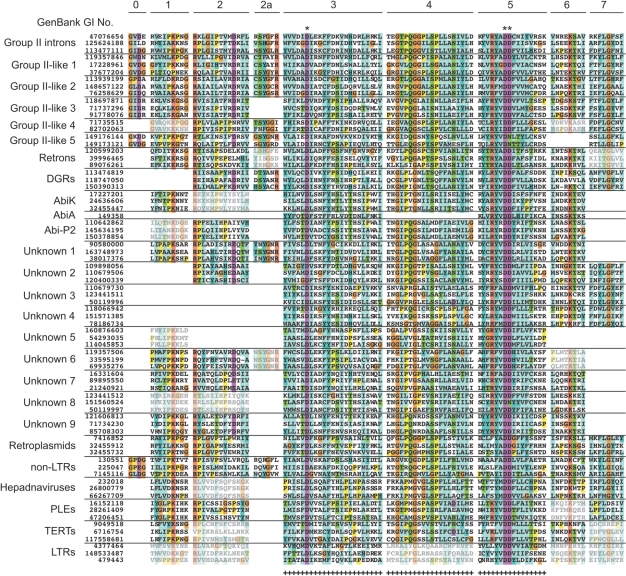
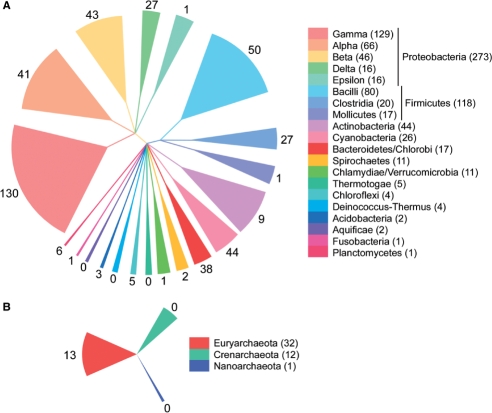
Retroelements are usually considered to be eukaryotic elements because of the large number and variety in eukaryotic genomes. By comparison, reverse transcriptases (RTs) are rare in bacteria, with only three characterized classes: retrons, group II introns and diversity-generating retroelements (DGRs). Here, we present the results of a bioinformatic survey that aims to define the landscape of RTs across eubacterial, archaeal and phage genomes. We identify and categorize 1021 RTs, of which the majority are group II introns (73%). Surprisingly, a plethora of novel RTs are found that do not belong to characterized classes. The RTs have 11 domain architectures and are classified into 20 groupings based on sequence similarity, phylogenetic analyses and open reading frame domain structures. Interestingly, group II introns are the only bacterial RTs to exhibit clear evidence for independent mobility, while five other groups have putative functions in defense against phage infection or promotion of phage infection. These examples suggest that additional beneficial functions will be discovered among uncharacterized RTs. The study lays the groundwork for experimental characterization of these highly diverse sequences and has implications for the evolution of retroelements.
Figures

References
-
- Baltimore D. RNA-dependent DNA polymerase in virions of RNA tumour viruses. Nature. 1970;226:1209–1211. - PubMed
-
- Temin HM, Mizutani S. RNA-dependent DNA polymerase in virions of Rous sarcoma virus. Nature. 1970;226:1211–1213. - PubMed
-
- Greider CW, Blackburn EH. A telomeric sequence in the RNA of Tetrahymena telomerase required for telomere repeat synthesis. Nature. 1989;337:331–337. - PubMed
-
- Eickbush TH. Origin and evolutionary relationships of retroelements. In: Morse SS, editor. The Evolutionary Biology of Viruses. New York: Raven Press; 1994. pp. 121–157.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
