

Optimization of human immunodeficiency virus gag expression by newcastle disease virus vectors for the induction of potent immune responses
- PMID: 19004953
- PMCID: PMC2612356
- DOI: 10.1128/JVI.01443-08
Optimization of human immunodeficiency virus gag expression by newcastle disease virus vectors for the induction of potent immune responses
Abstract
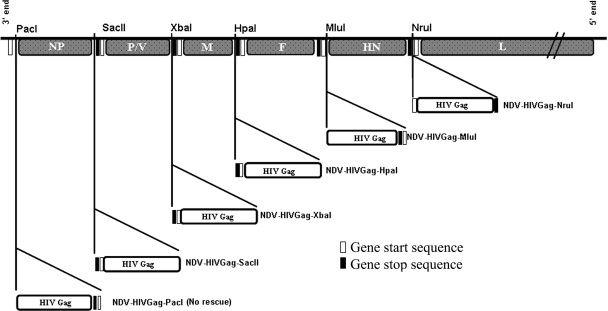
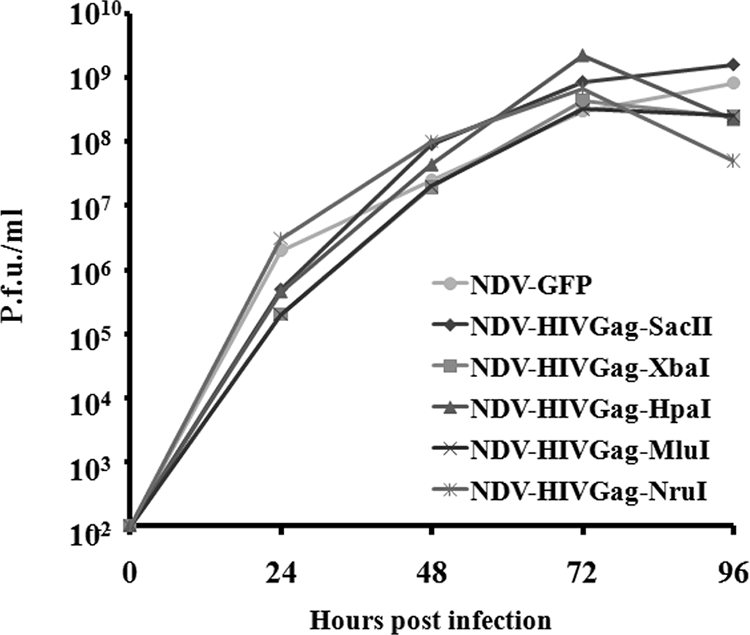
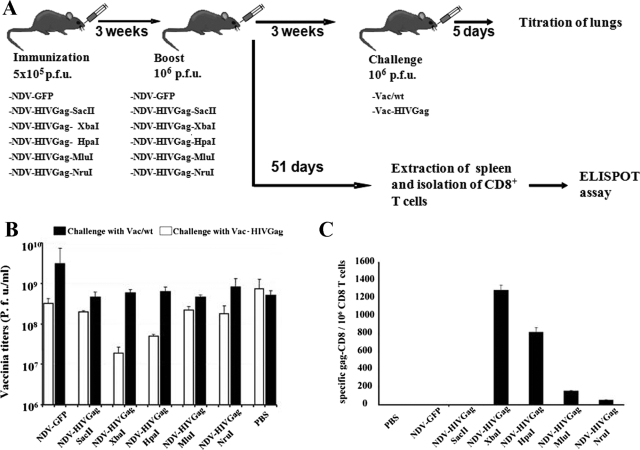
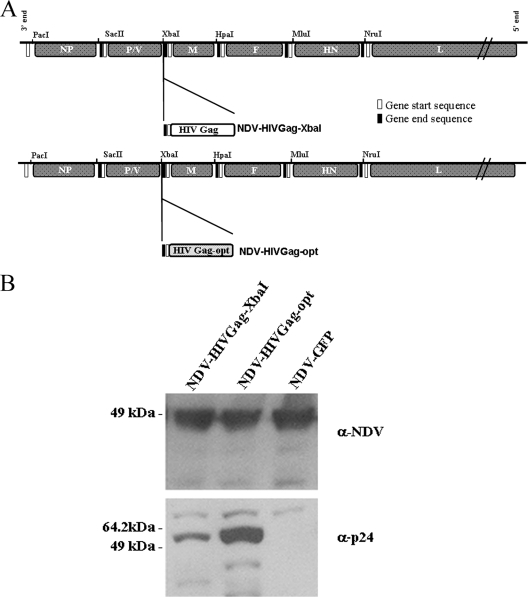
One attractive strategy for the development of a human immunodeficiency virus (HIV) vaccine is the use of viral vectors with a proven safety profile and an absence of preexisting immunity in humans, such as Newcastle disease virus (NDV). Several NDV vaccine vectors have been generated, and their immunogenicities have been investigated with different animal models. However, a systematic study to evaluate the optimal insertion site of the foreign antigens into NDV that results in enhanced immune responses specific to the antigen has not yet been conducted. In this article, we describe the ability of NDV expressing HIV Gag to generate a Gag-specific immune response in mice. We also have determined the optimal insertion site into the NDV genome by generating recombinant NDV-HIVGag viruses in which HIV gag was located at different transcriptional positions throughout the NDV viral genome. All recombinant viruses were viable, grew to similar titers in embryonated chicken eggs, and expressed Gag in a stable manner. Our in vivo experiments revealed that higher HIV Gag protein expression positively correlates with an enhanced CD8(+) T-cell-mediated immune response and protective immunity against challenge with vaccinia virus expressing HIV Gag. We also inserted a codon-optimized version of HIV gag in the described best location, between the P and M genes. Virus expressing the codon-optimized version of HIV gag induced a higher expression of the protein and an enhanced immune response against HIV Gag in mice. These results indicate that strategies directed toward increasing antigen expression by NDV result in enhanced immunogenicity and vaccine efficacy.
Figures
Similar articles
-
Mucosal Immunization with Newcastle Disease Virus Vector Coexpressing HIV-1 Env and Gag Proteins Elicits Potent Serum, Mucosal, and Cellular Immune Responses That Protect against Vaccinia Virus Env and Gag Challenges.mBio. 2015 Jul 21;6(4):e01005. doi: 10.1128/mBio.01005-15. mBio. 2015. PMID: 26199332 Free PMC article.
-
Evaluation of humoral, mucosal, and cellular immune responses following co-immunization of HIV-1 Gag and Env proteins expressed by Newcastle disease virus.Hum Vaccin Immunother. 2015;11(2):504-15. doi: 10.4161/21645515.2014.987006. Hum Vaccin Immunother. 2015. PMID: 25695657 Free PMC article.
-
Newcastle disease virus expressing a dendritic cell-targeted HIV gag protein induces a potent gag-specific immune response in mice.J Virol. 2011 Mar;85(5):2235-46. doi: 10.1128/JVI.02036-10. Epub 2010 Dec 15. J Virol. 2011. PMID: 21159873 Free PMC article.
-
Newcastle disease virus vectored vaccines as bivalent or antigen delivery vaccines.Clin Exp Vaccine Res. 2017 Jul;6(2):72-82. doi: 10.7774/cevr.2017.6.2.72. Epub 2017 Jul 26. Clin Exp Vaccine Res. 2017. PMID: 28775971 Free PMC article. Review.
-
Newcastle disease virus: a promising vector for viral therapy, immune therapy, and gene therapy of cancer.Methods Mol Biol. 2009;542:565-605. doi: 10.1007/978-1-59745-561-9_30. Methods Mol Biol. 2009. PMID: 19565923 Free PMC article. Review.
Cited by
-
Newcastle disease virus: current status and our understanding.Virus Res. 2014 May 12;184:71-81. doi: 10.1016/j.virusres.2014.02.016. Epub 2014 Mar 1. Virus Res. 2014. PMID: 24589707 Free PMC article. Review.
-
Newcastle Disease Virus as a Vaccine Vector for 20 Years: A Focus on Maternally Derived Antibody Interference.Vaccines (Basel). 2020 May 14;8(2):222. doi: 10.3390/vaccines8020222. Vaccines (Basel). 2020. PMID: 32422944 Free PMC article. Review.
-
Immunogenicity and protective efficacy of an intranasal live-attenuated vaccine against SARS-CoV-2.iScience. 2021 Sep 24;24(9):102941. doi: 10.1016/j.isci.2021.102941. Epub 2021 Aug 4. iScience. 2021. PMID: 34368648 Free PMC article.
-
Newcastle disease vaccines-A solved problem or a continuous challenge?Vet Microbiol. 2017 Jul;206:126-136. doi: 10.1016/j.vetmic.2016.12.019. Epub 2016 Dec 16. Vet Microbiol. 2017. PMID: 28024856 Free PMC article. Review.
-
Role of untranslated regions in regulation of gene expression, replication, and pathogenicity of Newcastle disease virus expressing green fluorescent protein.J Virol. 2010 Mar;84(5):2629-34. doi: 10.1128/JVI.02049-09. Epub 2009 Dec 16. J Virol. 2010. PMID: 20015997 Free PMC article.
References
-
- Addo, M. M., X. G. Yu, A. Rathod, D. Cohen, R. L. Eldridge, D. Strick, M. N. Johnston, C. Corcoran, A. G. Wurcel, C. A. Fitzpatrick, M. E. Feeney, W. R. Rodriguez, N. Basgoz, R. Draenert, D. R. Stone, C. Brander, P. J. Goulder, E. S. Rosenberg, M. Altfeld, and B. D. Walker. 2003. Comprehensive epitope analysis of human immunodeficiency virus type 1 (HIV-1)-specific T-cell responses directed against the entire expressed HIV-1 genome demonstrate broadly directed responses, but no correlation to viral load. J. Virol. 772081-2092. - PMC - PubMed
-
- Alexander, D. 1997. Newscastle disease and other avian Paramyxoviridae infections, p. 541-569. In B. W. Calnek et al. (ed.), Diseases of poultry, 10th ed. Iowa State University Press, Ames.
-
- Alexander, D. J. 2000. Newcastle disease and other avian paramyxoviruses. Rev. Sci. Tech. 19443-462. - PubMed
-
- Baba, T. W., V. Liska, A. H. Khimani, N. B. Ray, P. J. Dailey, D. Penninck, R. Bronson, M. F. Greene, H. M. McClure, L. N. Martin, and R. M. Ruprecht. 1999. Live attenuated, multiply deleted simian immunodeficiency virus causes AIDS in infant and adult macaques. Nat. Med. 5194-203. - PubMed
-
- Batliwalla, F. M., B. A. Bateman, D. Serrano, D. Murray, S. Macphail, V. C. Maino, J. C. Ansel, P. K. Gregersen, and C. A. Armstrong. 1998. A 15-year follow-up of AJCC stage III malignant melanoma patients treated postsurgically with Newcastle disease virus (NDV) oncolysate and determination of alterations in the CD8 T cell repertoire. Mol. Med. 4783-794. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
