

Adaptation of firing rate and spike-timing precision in the avian cochlear nucleus
- PMID: 19005056
- PMCID: PMC2693385
- DOI: 10.1523/JNEUROSCI.3827-08.2008
Adaptation of firing rate and spike-timing precision in the avian cochlear nucleus
Abstract
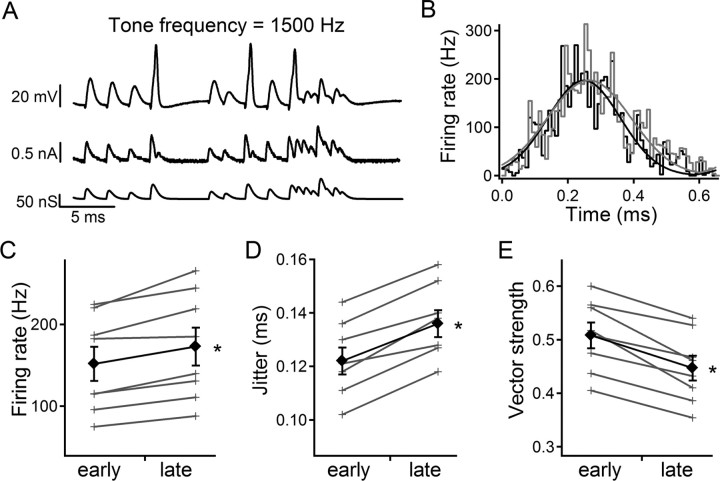
Adaptation is commonly defined as a decrease in response to a constant stimulus. In the auditory system such adaptation is seen at multiple levels. However, the first-order central neurons of the interaural time difference detection circuit encode information in the timing of spikes rather than the overall firing rate. We investigated adaptation during in vitro whole-cell recordings from chick nucleus magnocellularis neurons. Injection of noisy, depolarizing current caused an increase in firing rate and a decrease in spike time precision that developed over approximately 20 s. This adaptation depends on sustained depolarization, is independent of firing, and is eliminated by alpha-dendrotoxin (0.1 microM), implicating slow inactivation of low-threshold voltage-activated K+ channels as its mechanism. This process may alter both firing rate and spike-timing precision of phase-locked inputs to coincidence detector neurons in nucleus laminaris and thereby adjust the precision of sound localization.
Figures







Similar articles
-
Spike threshold adaptation diversifies neuronal operating modes in the auditory brain stem.J Neurophysiol. 2019 Dec 1;122(6):2576-2590. doi: 10.1152/jn.00234.2019. Epub 2019 Oct 2. J Neurophysiol. 2019. PMID: 31577531 Free PMC article.
-
Adaptation of spike timing precision controls the sensitivity to interaural time difference in the avian auditory brainstem.J Neurosci. 2012 Oct 31;32(44):15489-94. doi: 10.1523/JNEUROSCI.1865-12.2012. J Neurosci. 2012. PMID: 23115186 Free PMC article.
-
Early development of intrinsic and synaptic properties of chicken nucleus laminaris neurons.Neuroscience. 2008 Apr 22;153(1):131-43. doi: 10.1016/j.neuroscience.2008.01.059. Epub 2008 Feb 13. Neuroscience. 2008. PMID: 18355968
-
Beyond timing in the auditory brainstem: intensity coding in the avian cochlear nucleus angularis.Prog Brain Res. 2007;165:123-33. doi: 10.1016/S0079-6123(06)65008-5. Prog Brain Res. 2007. PMID: 17925243 Free PMC article. Review.
-
Neuronal specializations for the processing of interaural difference cues in the chick.Front Neural Circuits. 2014 May 9;8:47. doi: 10.3389/fncir.2014.00047. eCollection 2014. Front Neural Circuits. 2014. PMID: 24847212 Free PMC article. Review.
Cited by
-
Spike threshold adaptation diversifies neuronal operating modes in the auditory brain stem.J Neurophysiol. 2019 Dec 1;122(6):2576-2590. doi: 10.1152/jn.00234.2019. Epub 2019 Oct 2. J Neurophysiol. 2019. PMID: 31577531 Free PMC article.
-
Coding strategies in the otolith system differ for translational head motion vs. static orientation relative to gravity.Elife. 2019 Jun 14;8:e45573. doi: 10.7554/eLife.45573. Elife. 2019. PMID: 31199243 Free PMC article.
-
Adaptation of spike timing precision controls the sensitivity to interaural time difference in the avian auditory brainstem.J Neurosci. 2012 Oct 31;32(44):15489-94. doi: 10.1523/JNEUROSCI.1865-12.2012. J Neurosci. 2012. PMID: 23115186 Free PMC article.
-
Spike timing precision changes with spike rate adaptation in the owl's auditory space map.J Neurophysiol. 2015 Oct;114(4):2204-19. doi: 10.1152/jn.00442.2015. Epub 2015 Aug 12. J Neurophysiol. 2015. PMID: 26269555 Free PMC article.
-
Self-motion evokes precise spike timing in the primate vestibular system.Nat Commun. 2016 Oct 27;7:13229. doi: 10.1038/ncomms13229. Nat Commun. 2016. PMID: 27786265 Free PMC article.
References
-
- Bacci A, Huguenard JR. Enhancement of spike-timing precision by autaptic transmission in neocortical inhibitory interneurons. Neuron. 2006;49:119–130. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources