

The UNI1 and UNI2 genes function in the transition of triplet to doublet microtubules between the centriole and cilium in Chlamydomonas
- PMID: 19005206
- PMCID: PMC2613092
- DOI: 10.1091/mbc.e08-09-0900
The UNI1 and UNI2 genes function in the transition of triplet to doublet microtubules between the centriole and cilium in Chlamydomonas
Abstract
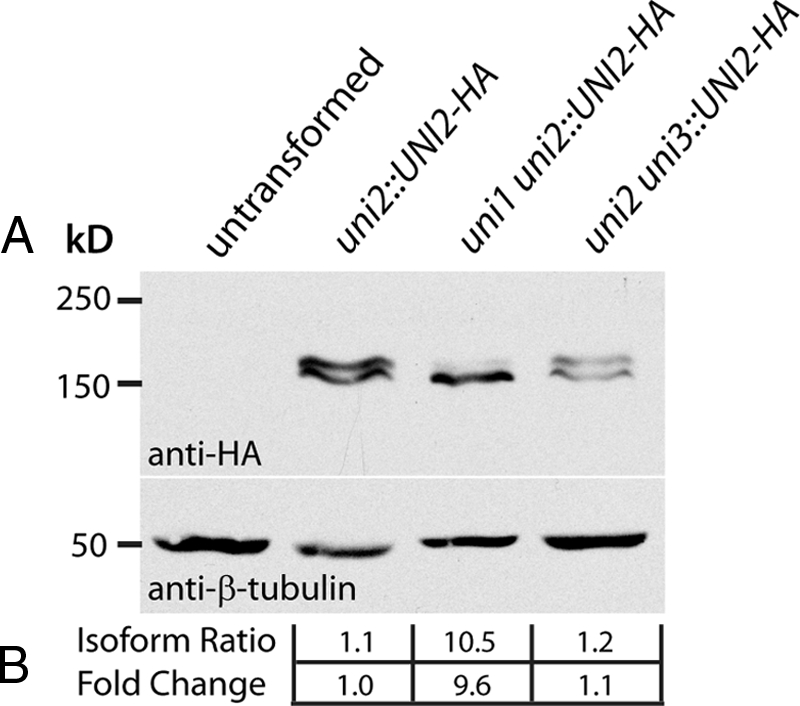
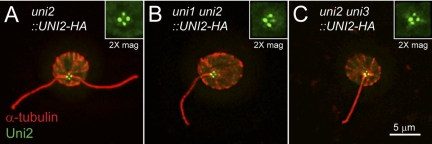
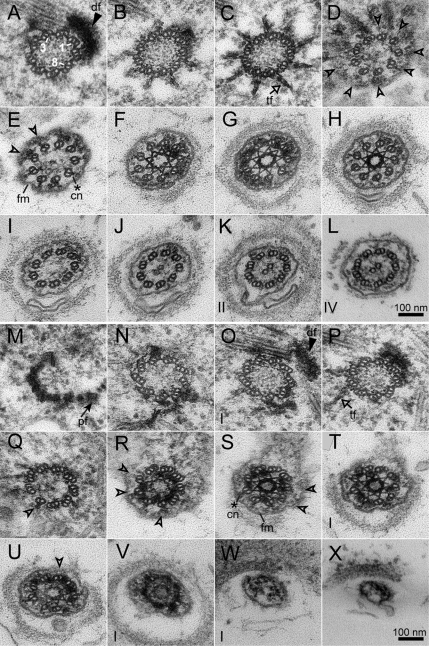
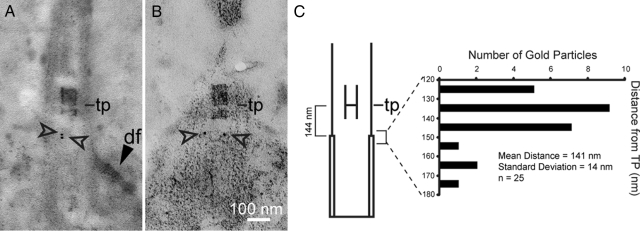
One fundamental role of the centriole in eukaryotic cells is to nucleate the growth of cilia. The unicellular alga Chlamydomonas reinhardtii provides a simple genetic system to study the role of the centriole in ciliogenesis. Wild-type cells are biflagellate, but "uni" mutations result in failure of some centrioles (basal bodies) to assemble cilia (flagella). Serial transverse sections through basal bodies in uni1 and uni2 single and double mutant cells revealed a previously undescribed defect in the transition of triplet microtubules to doublet microtubules, a defect correlated with failure to assemble flagella. Phosphorylation of the Uni2 protein is reduced in uni1 mutant cells. Immunogold electron microscopy showed that the Uni2 protein localizes at the distal end of the basal body where microtubule transition occurs. These results provide the first mechanistic insights into the function of UNI1 and UNI2 genes in the pathway mediating assembly of doublet microtubules in the axoneme from triplet microtubules in the basal body template.
Figures


Similar articles
-
Bld10p, a novel protein essential for basal body assembly in Chlamydomonas: localization to the cartwheel, the first ninefold symmetrical structure appearing during assembly.J Cell Biol. 2004 Jun 7;165(5):663-71. doi: 10.1083/jcb.200402022. Epub 2004 Jun 1. J Cell Biol. 2004. PMID: 15173189 Free PMC article.
-
Three-dimensional organization of basal bodies from wild-type and delta-tubulin deletion strains of Chlamydomonas reinhardtii.Mol Biol Cell. 2003 Jul;14(7):2999-3012. doi: 10.1091/mbc.e02-11-0755. Epub 2003 Apr 4. Mol Biol Cell. 2003. PMID: 12857881 Free PMC article.
-
Katanin localization requires triplet microtubules in Chlamydomonas reinhardtii.PLoS One. 2013;8(1):e53940. doi: 10.1371/journal.pone.0053940. Epub 2013 Jan 8. PLoS One. 2013. PMID: 23320108 Free PMC article.
-
Elucidation of basal body and centriole functions in Chlamydomonas reinhardtii.Traffic. 2003 Jul;4(7):443-51. doi: 10.1034/j.1600-0854.2003.00104.x. Traffic. 2003. PMID: 12795689 Review.
-
Microtubule Inner Proteins: A Meshwork of Luminal Proteins Stabilizing the Doublet Microtubule.Bioessays. 2018 Mar;40(3). doi: 10.1002/bies.201700209. Epub 2018 Feb 12. Bioessays. 2018. PMID: 29430673 Review.
Cited by
-
TIM, a targeted insertional mutagenesis method utilizing CRISPR/Cas9 in Chlamydomonas reinhardtii.PLoS One. 2020 May 13;15(5):e0232594. doi: 10.1371/journal.pone.0232594. eCollection 2020. PLoS One. 2020. PMID: 32401787 Free PMC article.
-
Cep120 is asymmetrically localized to the daughter centriole and is essential for centriole assembly.J Cell Biol. 2010 Oct 18;191(2):331-46. doi: 10.1083/jcb.201003009. J Cell Biol. 2010. PMID: 20956381 Free PMC article.
-
Asymmetric properties of the Chlamydomonas reinhardtii cytoskeleton direct rhodopsin photoreceptor localization.J Cell Biol. 2011 May 16;193(4):741-53. doi: 10.1083/jcb.201009131. Epub 2011 May 9. J Cell Biol. 2011. PMID: 21555459 Free PMC article.
-
CCDC61/VFL3 Is a Paralog of SAS6 and Promotes Ciliary Functions.Structure. 2020 Jun 2;28(6):674-689.e11. doi: 10.1016/j.str.2020.04.010. Epub 2020 May 5. Structure. 2020. PMID: 32375023 Free PMC article.
-
Genetic and genomic approaches to identify genes involved in flagellar assembly in Chlamydomonas reinhardtii.Methods Cell Biol. 2015;127:349-86. doi: 10.1016/bs.mcb.2014.12.001. Epub 2015 Feb 14. Methods Cell Biol. 2015. PMID: 25837400 Free PMC article.
References
-
- Albrecht-Buehler G., Bushnell A. The ultrastructure of primary cilia in quiescent 3T3 cells. Exp. Cell Res. 1980;126:427–437. - PubMed
-
- Beech P. L., Heimann K., Melkonian M. Development of the flagellar apparatus during the cell cycle in unicellular algae. Protoplasma. 1991;164:23–37.
-
- Cavalier-Smith T. Basal body and flagellar development during the vegetative cell cycle and the sexual cycle of Chlamydomonas reinhardtii. J. Cell Sci. 1974;16:529–556. - PubMed
-
- Cavalier-Smith T. The phagotrophic origin of eukaryotes and phylogenetic classification of Protozoa. Int. J. Syst. Evol. Microbiol. 2002;52:297–354. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
