

Axospinous synaptic subtype-specific differences in structure, size, ionotropic receptor expression, and connectivity in apical dendritic regions of rat hippocampal CA1 pyramidal neurons
- PMID: 19006199
- PMCID: PMC2592507
- DOI: 10.1002/cne.21896
Axospinous synaptic subtype-specific differences in structure, size, ionotropic receptor expression, and connectivity in apical dendritic regions of rat hippocampal CA1 pyramidal neurons
Abstract
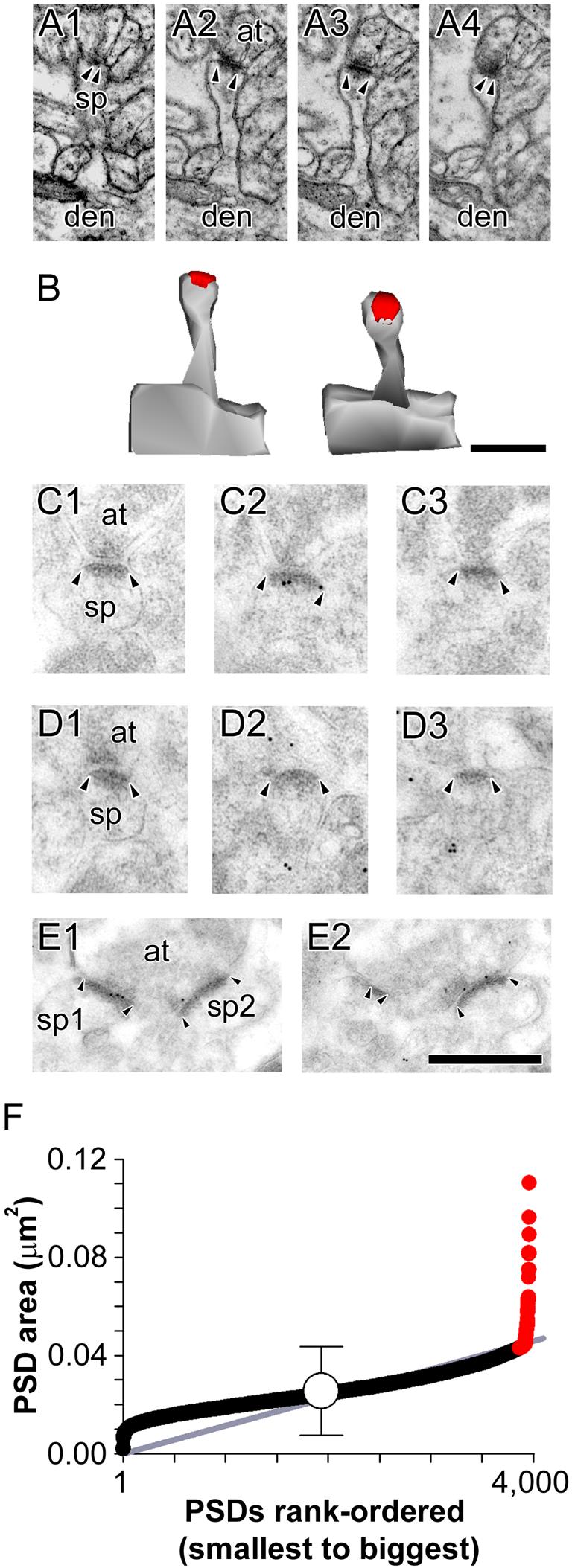
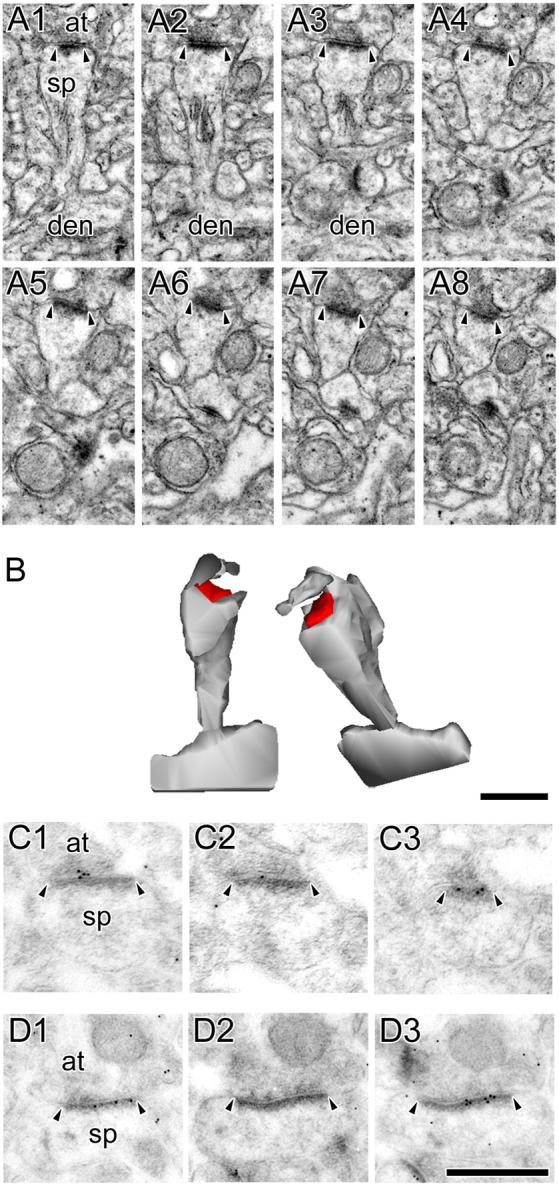
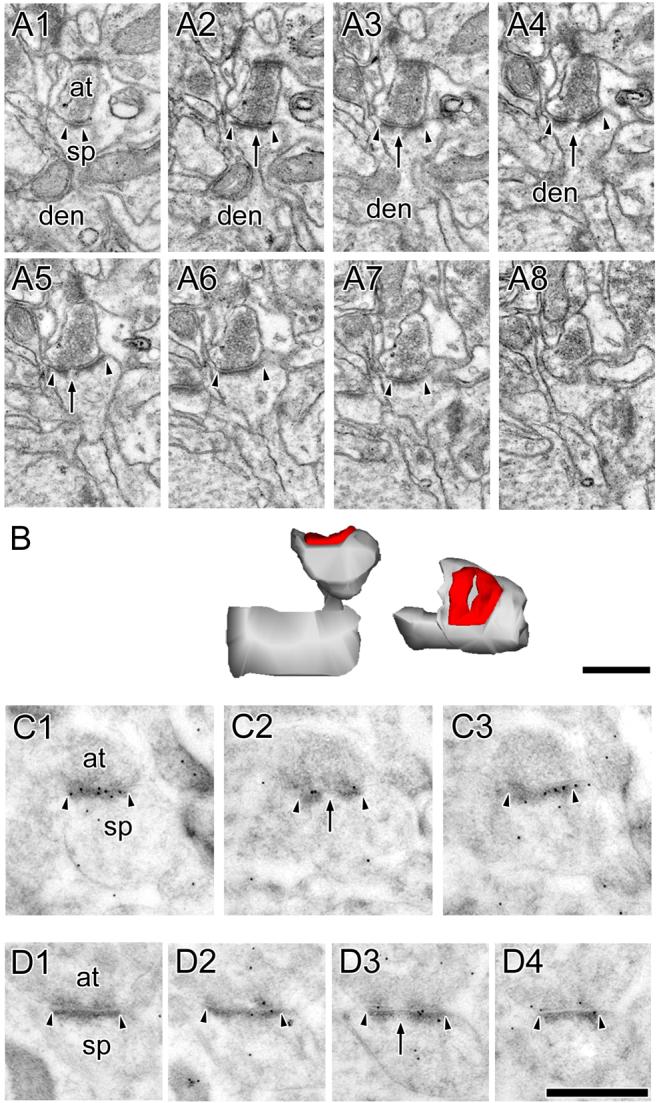
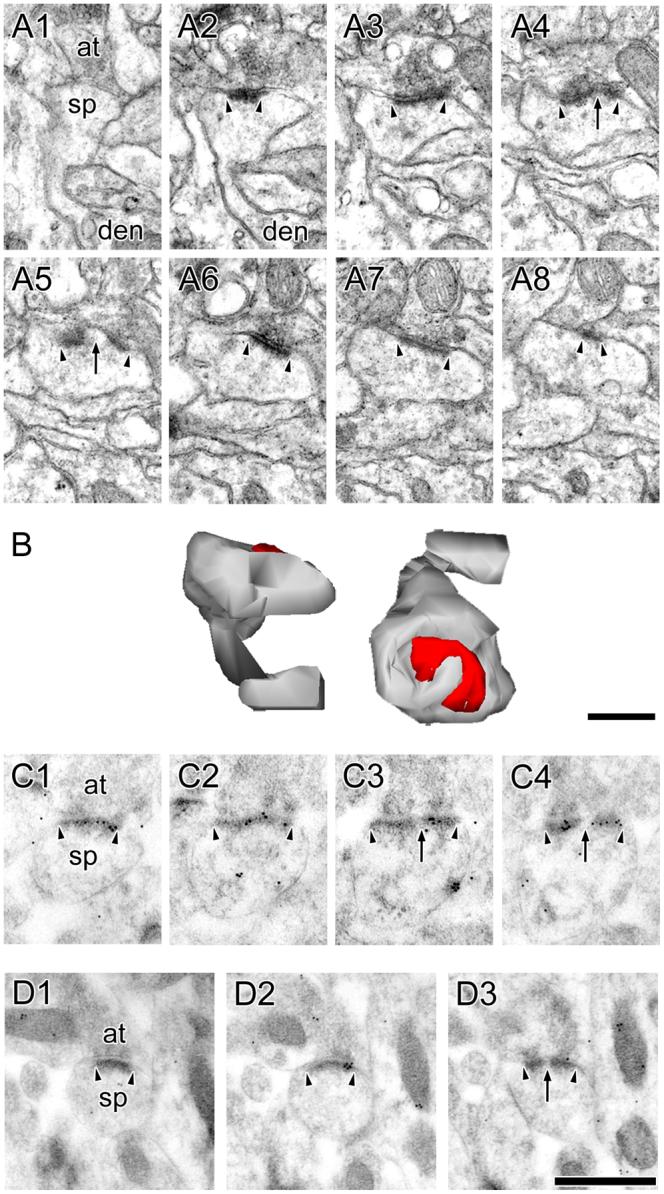
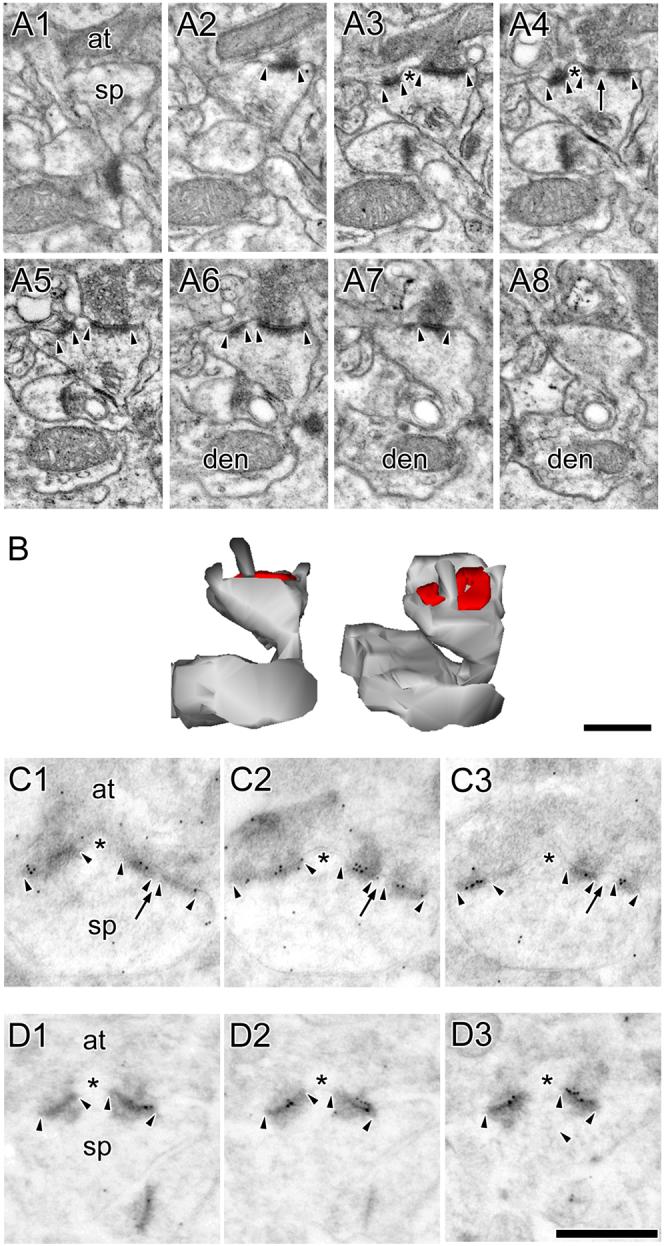
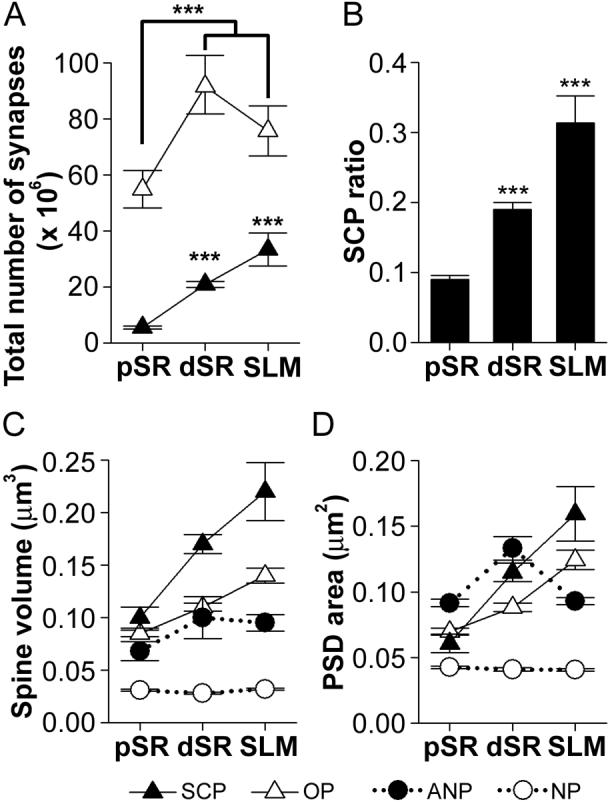
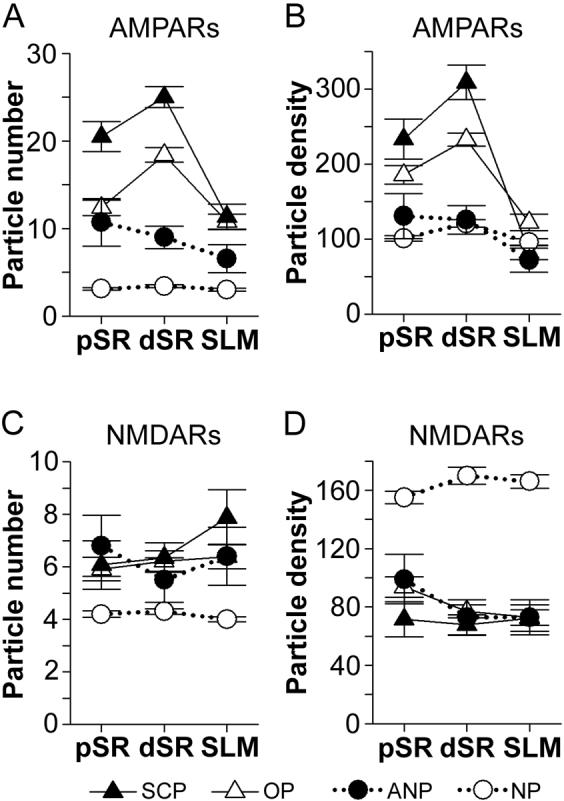
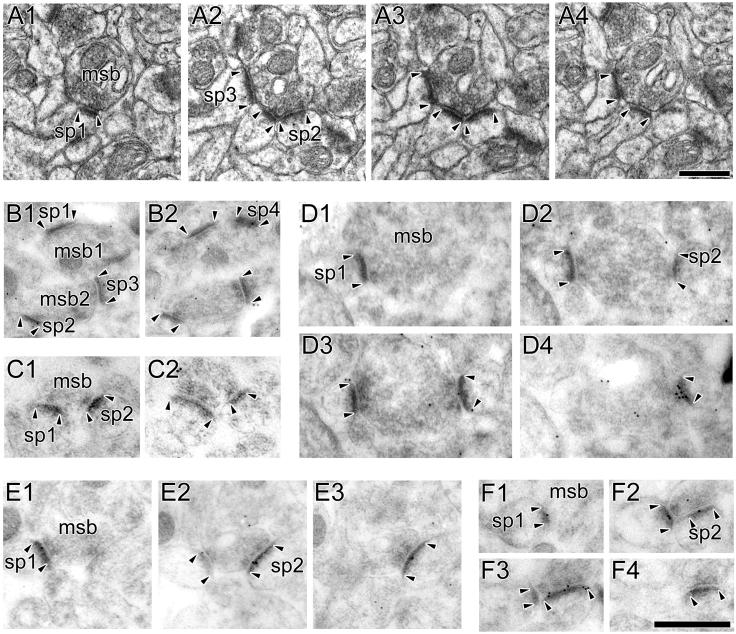
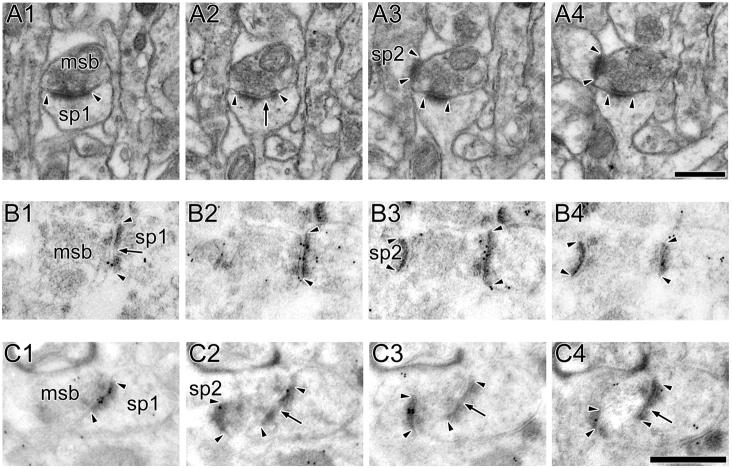
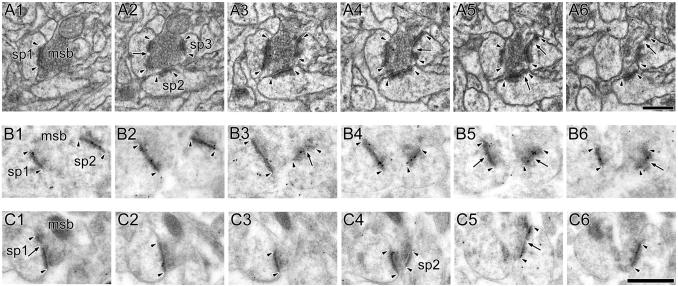
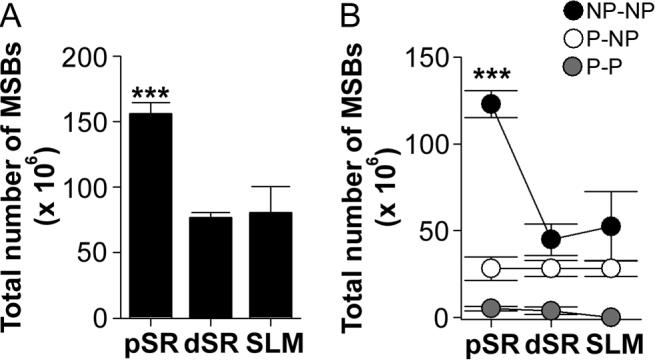
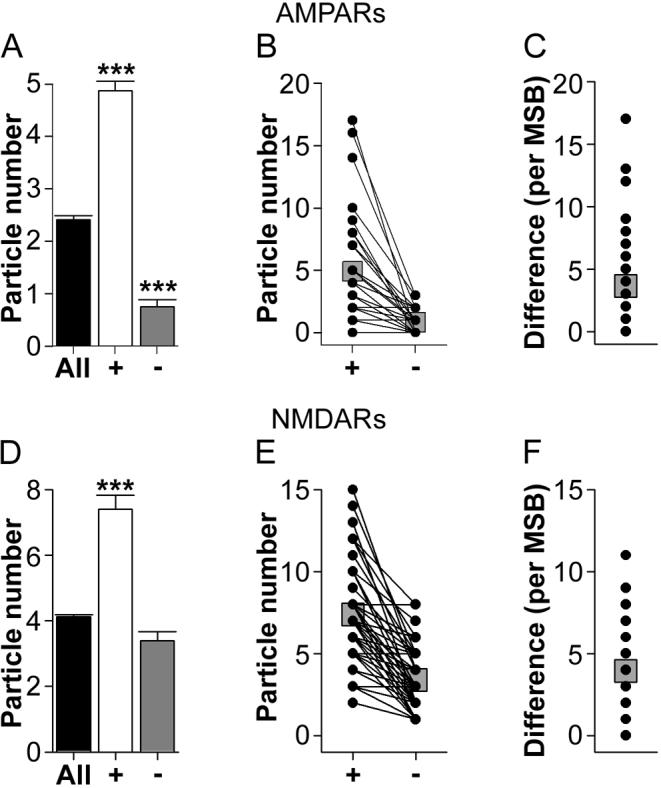
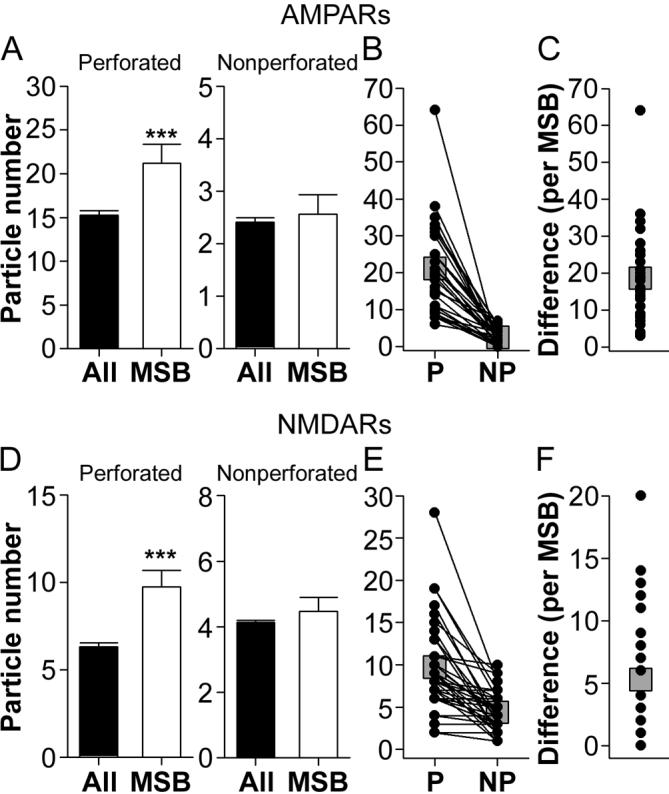
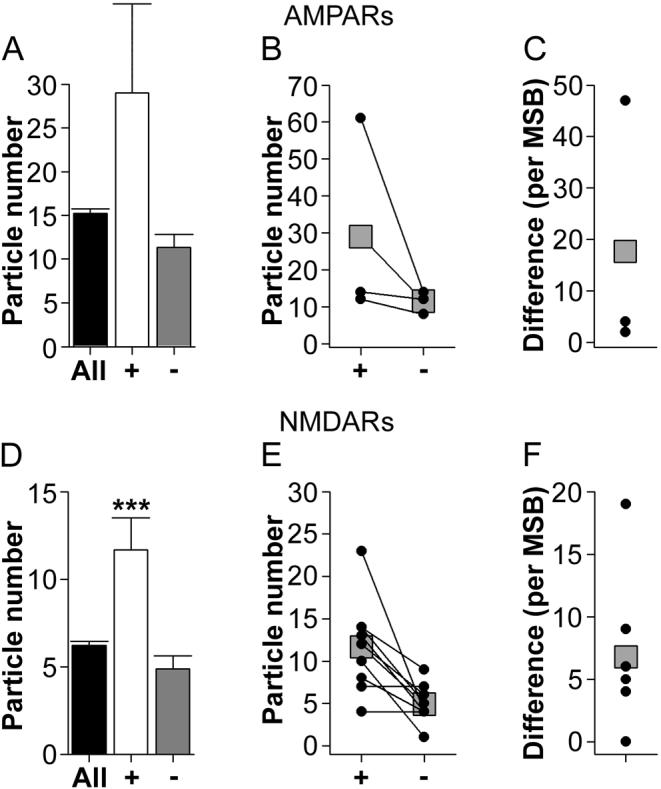
The morphology of axospinous synapses and their parent spines varies widely. Additionally, many of these synapses are contacted by multiple synapse boutons (MSBs) and show substantial variability in receptor expression. The two major axospinous synaptic subtypes are perforated and nonperforated, but there are several subcategories within these two classes. The present study used serial section electron microscopy to determine whether perforated and nonperforated synaptic subtypes differed with regard to their distribution, size, receptor expression, and connectivity to MSBs in three apical dendritic regions of rat hippocampal area CA1: the proximal and distal thirds of stratum radiatum, and the stratum lacunosum-moleculare. All synaptic subtypes were present throughout the apical dendritic regions, but there were several subclass-specific differences. First, segmented, completely partitioned synapses changed in number, proportion, and alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionate (AMPA) receptor expression with distance from the soma beyond that found within other perforated synaptic subtypes. Second, atypically large, nonperforated synapses showed N-methyl-D-aspartate (NMDA) receptor immunoreactivity identical to that of perforated synapses, levels of AMPA receptor expression intermediate to that of nonperforated and perforated synapses, and perforated synapse-like changes in structure with distance from the soma. Finally, MSB connectivity was highest in the proximal stratum radiatum, but only for those MSBs composed of nonperforated synapses. The immunogold data suggest that most MSBs would not generate simultaneous depolarizations in multiple neurons or spines, however, because the vast majority of MSBs are comprised of two synapses with abnormally low levels of receptor expression, or involve one synapse with a high level of receptor expression and another with only a low level.
Figures
Similar articles
-
Distance-dependent differences in synapse number and AMPA receptor expression in hippocampal CA1 pyramidal neurons.Neuron. 2006 May 4;50(3):431-42. doi: 10.1016/j.neuron.2006.03.022. Neuron. 2006. PMID: 16675397
-
Differences in the expression of AMPA and NMDA receptors between axospinous perforated and nonperforated synapses are related to the configuration and size of postsynaptic densities.J Comp Neurol. 2004 Jan 1;468(1):86-95. doi: 10.1002/cne.10950. J Comp Neurol. 2004. PMID: 14648692
-
Synapses with a segmented, completely partitioned postsynaptic density express more AMPA receptors than other axospinous synaptic junctions.Neuroscience. 2004;125(3):615-23. doi: 10.1016/j.neuroscience.2004.02.025. Neuroscience. 2004. PMID: 15099675
-
Large variability in synaptic N-methyl-D-aspartate receptor density on interneurons and a comparison with pyramidal-cell spines in the rat hippocampus.Neuroscience. 2003;119(2):347-63. doi: 10.1016/s0306-4522(03)00157-x. Neuroscience. 2003. PMID: 12770551
-
Synapse distribution suggests a two-stage model of dendritic integration in CA1 pyramidal neurons.Neuron. 2009 Jul 30;63(2):171-7. doi: 10.1016/j.neuron.2009.06.023. Neuron. 2009. PMID: 19640476 Free PMC article. Review.
Cited by
-
Effects of sleep and waking on the synaptic ultrastructure.Philos Trans R Soc Lond B Biol Sci. 2020 May 25;375(1799):20190235. doi: 10.1098/rstb.2019.0235. Epub 2020 Apr 6. Philos Trans R Soc Lond B Biol Sci. 2020. PMID: 32248785 Free PMC article.
-
Optogenetics stimulates nerve reorganization in the contralesional anterolateral primary motor cortex in a mouse model of ischemic stroke.Neural Regen Res. 2022 Jul;17(7):1535-1544. doi: 10.4103/1673-5374.330615. Neural Regen Res. 2022. PMID: 34916439 Free PMC article.
-
Multi-synaptic boutons are a feature of CA1 hippocampal connections in the stratum oriens.Cell Rep. 2023 May 30;42(5):112397. doi: 10.1016/j.celrep.2023.112397. Epub 2023 Apr 18. Cell Rep. 2023. PMID: 37074915 Free PMC article.
-
Physiological and anatomical studies of associative learning: Convergence with learning studies of W.T. Greenough.Dev Psychobiol. 2011 Jul;53(5):489-504. doi: 10.1002/dev.20554. Dev Psychobiol. 2011. PMID: 21678397 Free PMC article. Review.
-
Immunogold electron microscopic evidence of differential regulation of GluN1, GluN2A, and GluN2B, NMDA-type glutamate receptor subunits in rat hippocampal CA1 synapses during benzodiazepine withdrawal.J Comp Neurol. 2010 Nov 1;518(21):4311-28. doi: 10.1002/cne.22458. J Comp Neurol. 2010. PMID: 20853509 Free PMC article.
References
-
- Alvarez VA, Sabatini BL. Anatomical and physiological plasticity of dendritic spines. Annu Rev Neurosci. 2007;30:79–97. - PubMed
-
- Bloodgood BL, Sabatini BL. Neuronal activity regulates diffusion across the neck of dendritic spines. Science. 2005;310:866–869. - PubMed
-
- Bourne J, Harris KM. Do thin spines learn to be mushroom spines that remember? Curr Opin Neurobiol. 2007;17:381–386. - PubMed
-
- Bredt DS, Nicoll RA. AMPA receptor trafficking at excitatory synapses. Neuron. 2003;40:361–379. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
