

Products and substrate/template usage of vaccinia virus DNA primase
- PMID: 19007959
- PMCID: PMC2630392
- DOI: 10.1016/j.virol.2008.10.008
Products and substrate/template usage of vaccinia virus DNA primase
Abstract
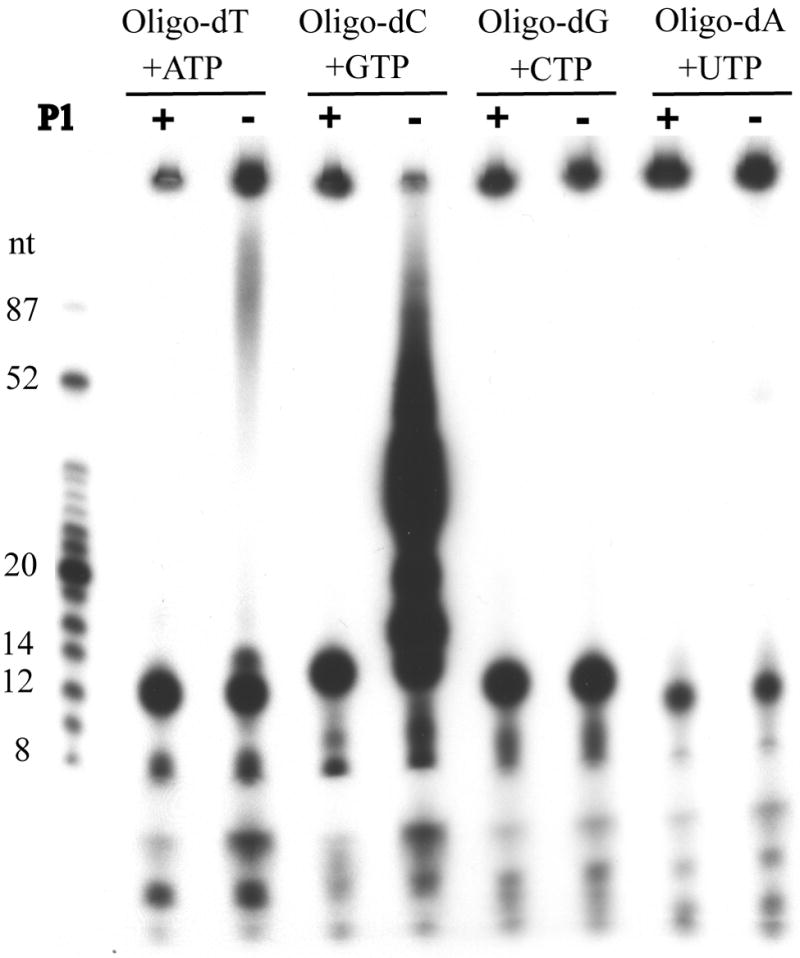
Vaccinia virus encodes a 90-kDa protein conserved in all poxviruses, with DNA primase and nucleoside triphosphatase activities. DNA primase products, synthesized with a single stranded varphiX174 DNA template, were resolved as dinucleotides and long RNAs on denaturing polyacrylamide and agarose gels. Following phosphatase treatment, the dinucleotides GpC and ApC in a 4:1 ratio were identified by nearest neighbor analysis in which (32)P was transferred from [alpha-(32)P]CTP to initiating purine nucleotides. Differences in the nucleotide binding sites for initiation and elongation were suggested by the absence of CpC and UpC dinucleotides as well as the inability of deoxynucleotides to mediate primer synthesis despite their incorporation into mixed RNA/DNA primers. Strong primase activity was detected with an oligo(dC) template. However, there was only weak activity with an oligo(dT) template and none with oligo(dA) or oligo(dG). The absence of stringent template specificity is consistent with a role for the enzyme in priming DNA synthesis at the replication fork.
Figures
Similar articles
-
Domain Organization of Vaccinia Virus Helicase-Primase D5.J Virol. 2016 Apr 14;90(9):4604-4613. doi: 10.1128/JVI.00044-16. Print 2016 May. J Virol. 2016. PMID: 26912611 Free PMC article.
-
RNA priming coupled with DNA synthesis on natural template by calf thymus DNA polymerase alpha-primase.Biochemistry. 1993 Nov 30;32(47):12782-92. doi: 10.1021/bi00210a030. Biochemistry. 1993. PMID: 7504526
-
Key role of template sequence for primer synthesis by the herpes simplex virus 1 helicase-primase.Biochemistry. 2002 Dec 10;41(49):14569-79. doi: 10.1021/bi026680v. Biochemistry. 2002. PMID: 12463757
-
Interaction of ribonucleoside triphosphates with the gene 4 primase of bacteriophage T7.J Biol Chem. 1999 Dec 10;274(50):35899-907. doi: 10.1074/jbc.274.50.35899. J Biol Chem. 1999. PMID: 10585475
-
Initiation of DNA synthesis by the calf thymus DNA polymerase-primase complex.J Biol Chem. 1985 Sep 5;260(19):10840-6. J Biol Chem. 1985. PMID: 3161883
Cited by
-
Low-resolution structure of vaccinia virus DNA replication machinery.J Virol. 2013 Feb;87(3):1679-89. doi: 10.1128/JVI.01533-12. Epub 2012 Nov 21. J Virol. 2013. PMID: 23175373 Free PMC article.
-
Characterization of indels in poxvirus genomes.Virus Genes. 2011 Apr;42(2):171-7. doi: 10.1007/s11262-010-0560-x. Epub 2010 Dec 14. Virus Genes. 2011. PMID: 21153876
-
Primase-polymerases are a functionally diverse superfamily of replication and repair enzymes.Nucleic Acids Res. 2015 Aug 18;43(14):6651-64. doi: 10.1093/nar/gkv625. Epub 2015 Jun 24. Nucleic Acids Res. 2015. PMID: 26109351 Free PMC article. Review.
-
Mimivirus encodes a multifunctional primase with DNA/RNA polymerase, terminal transferase and translesion synthesis activities.Nucleic Acids Res. 2019 Jul 26;47(13):6932-6945. doi: 10.1093/nar/gkz236. Nucleic Acids Res. 2019. PMID: 31001622 Free PMC article.
-
Cellular DNA ligase I is recruited to cytoplasmic vaccinia virus factories and masks the role of the vaccinia ligase in viral DNA replication.Cell Host Microbe. 2009 Dec 17;6(6):563-9. doi: 10.1016/j.chom.2009.11.005. Cell Host Microbe. 2009. PMID: 20006844 Free PMC article.
References
-
- Baroudy BM, Venkatesan S, Moss B. Incompletely base-paired flipflop terminal loops link the two DNA strands of the vaccinia virus genome into one uninterrupted polynucleotide chain. Cell. 1982a;28:315–324. - PubMed
-
- Baroudy BM, Venkatesan S, Moss B. Structure and replication of vaccinia virus telomeres. Cold Spring Harbor Symp Quant Biol. 1982b;47:723–729. - PubMed
-
- Bocquier AA, Liu L, Cann IK, Komori K, Kohda D, Ishino Y. Archaeal primase: bridging the gap between RNA and DNA polymerases. Curr Biol. 2001;11:452–456. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
