

Redox modification of ryanodine receptors contributes to sarcoplasmic reticulum Ca2+ leak in chronic heart failure
- PMID: 19008475
- PMCID: PMC3274754
- DOI: 10.1161/CIRCRESAHA.108.184457
Redox modification of ryanodine receptors contributes to sarcoplasmic reticulum Ca2+ leak in chronic heart failure
Abstract
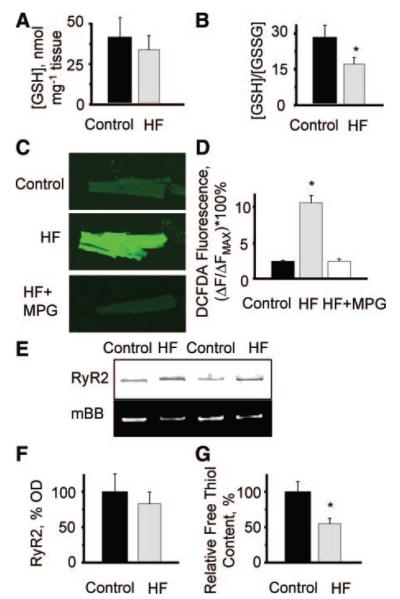
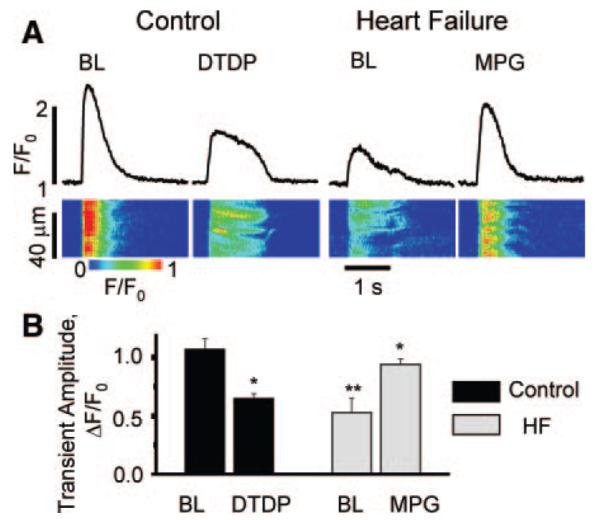
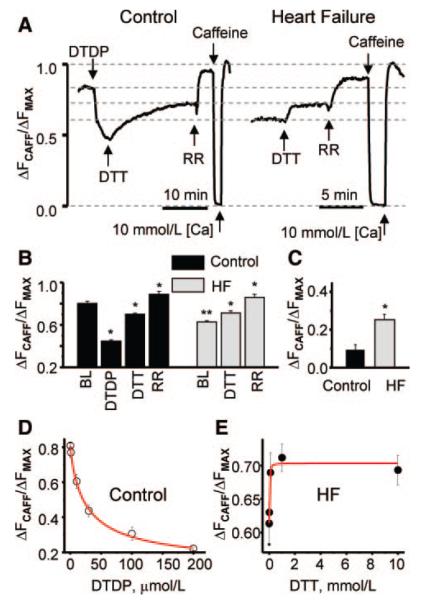
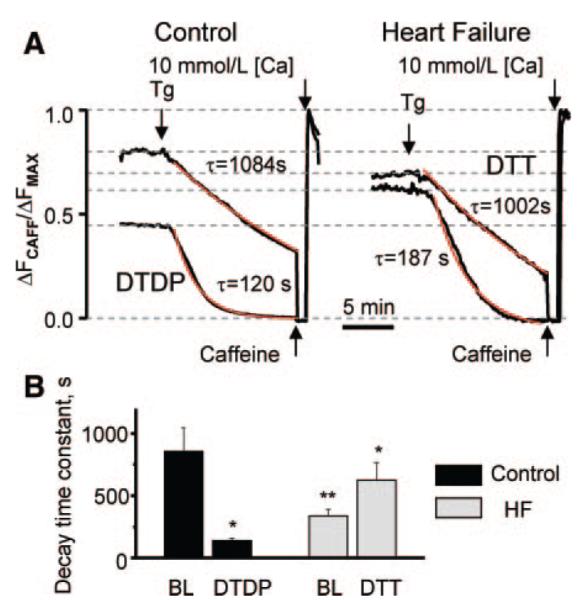
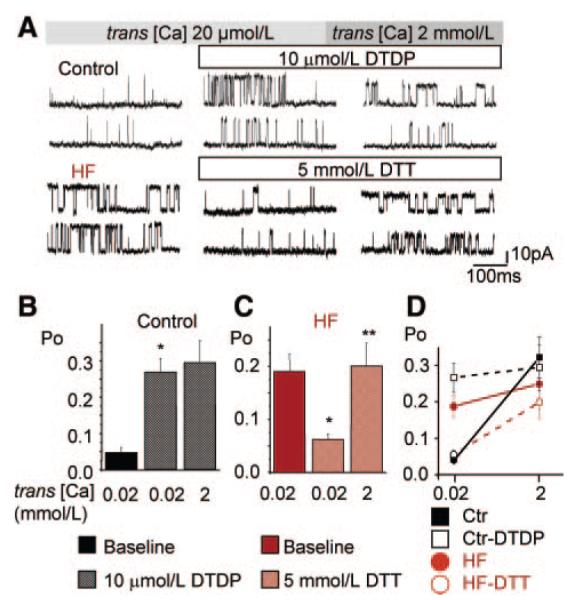
Abnormal cardiac ryanodine receptor (RyR2) function is recognized as an important factor in the pathogenesis of heart failure (HF). However, the specific molecular causes underlying RyR2 defects in HF remain poorly understood. In the present study, we used a canine model of chronic HF to test the hypothesis that the HF-related alterations in RyR2 function are caused by posttranslational modification by reactive oxygen species generated in the failing heart. Experimental approaches included imaging of cytosolic ([Ca(2+)](c)) and sarcoplasmic reticulum (SR) luminal Ca(2+) ([Ca(2+)]SR) in isolated intact and permeabilized ventricular myocytes and single RyR2 channel recording using the planar lipid bilayer technique. The ratio of reduced to oxidized glutathione, as well as the level of free thiols on RyR2 decreased markedly in failing versus control hearts consistent with increased oxidative stress in HF. RyR2-mediated SR Ca(2+) leak was significantly enhanced in permeabilized myocytes, resulting in reduced [Ca(2+)](SR) in HF compared to control cells. Both SR Ca(2+) leak and [Ca(2+)](SR) were partially normalized by treating HF myocytes with reducing agents. Conversely, oxidizing agents accelerated SR Ca(2+) leak and decreased [Ca(2+)](SR) in cells from normal hearts. Moreover, exposure to antioxidants significantly improved intracellular Ca(2+)-handling parameters in intact HF myocytes. Single RyR2 channel activity was significantly higher in HF versus control because of increased sensitivity to activation by luminal Ca(2+) and was partially normalized by reducing agents through restoring luminal Ca(2+) sensitivity oxidation of control RyR2s enhanced their luminal Ca(2+) sensitivity, thus reproducing the HF phenotype. These findings suggest that redox modification contributes to abnormal function of RyR2s in HF, presenting a potential therapeutic target for treating HF.
Figures
Similar articles
-
Enhanced ryanodine receptor-mediated calcium leak determines reduced sarcoplasmic reticulum calcium content in chronic canine heart failure.Biophys J. 2007 Dec 1;93(11):4083-92. doi: 10.1529/biophysj.107.114546. Epub 2007 Sep 7. Biophys J. 2007. PMID: 17827226 Free PMC article.
-
Ca2+/calmodulin-dependent protein kinase modulates cardiac ryanodine receptor phosphorylation and sarcoplasmic reticulum Ca2+ leak in heart failure.Circ Res. 2005 Dec 9;97(12):1314-22. doi: 10.1161/01.RES.0000194329.41863.89. Epub 2005 Nov 3. Circ Res. 2005. PMID: 16269653
-
The role of RyR2 oxidation in the blunted frequency-dependent facilitation of Ca2+ transient amplitude in rabbit failing myocytes.Pflugers Arch. 2018 Jun;470(6):959-968. doi: 10.1007/s00424-018-2122-3. Epub 2018 Mar 2. Pflugers Arch. 2018. PMID: 29500669 Free PMC article.
-
Intracellular calcium leak in heart failure and atrial fibrillation: a unifying mechanism and therapeutic target.Nat Rev Cardiol. 2020 Nov;17(11):732-747. doi: 10.1038/s41569-020-0394-8. Epub 2020 Jun 18. Nat Rev Cardiol. 2020. PMID: 32555383 Free PMC article. Review.
-
Functional Impact of Ryanodine Receptor Oxidation on Intracellular Calcium Regulation in the Heart.Rev Physiol Biochem Pharmacol. 2016;171:39-62. doi: 10.1007/112_2016_2. Rev Physiol Biochem Pharmacol. 2016. PMID: 27251471 Free PMC article. Review.
Cited by
-
Single-Cell Mechanical Analysis of Human Pluripotent Stem Cell-Derived Cardiomyocytes for Drug Testing and Pathophysiological Studies.Stem Cell Reports. 2020 Sep 8;15(3):587-596. doi: 10.1016/j.stemcr.2020.07.006. Epub 2020 Aug 6. Stem Cell Reports. 2020. PMID: 32763158 Free PMC article.
-
Sarcoplasmic reticulum-mitochondria communication; implications for cardiac arrhythmia.J Mol Cell Cardiol. 2021 Jul;156:105-113. doi: 10.1016/j.yjmcc.2021.04.002. Epub 2021 Apr 17. J Mol Cell Cardiol. 2021. PMID: 33857485 Free PMC article. Review.
-
Heart failure duration progressively modulates the arrhythmia substrate through structural and electrical remodeling.Life Sci. 2015 Feb 15;123:61-71. doi: 10.1016/j.lfs.2014.12.024. Epub 2015 Jan 14. Life Sci. 2015. PMID: 25596015 Free PMC article.
-
Enhancing Autophagy Diminishes Aberrant Ca2+ Homeostasis and Arrhythmogenesis in Aging Rabbit Hearts.Front Physiol. 2019 Oct 4;10:1277. doi: 10.3389/fphys.2019.01277. eCollection 2019. Front Physiol. 2019. PMID: 31636573 Free PMC article.
-
Alteration of sarcoplasmic reticulum Ca2+ release termination by ryanodine receptor sensitization and in heart failure.J Physiol. 2009 Nov 1;587(Pt 21):5197-209. doi: 10.1113/jphysiol.2009.177576. Epub 2009 Sep 7. J Physiol. 2009. PMID: 19736296 Free PMC article.
References
-
- Lindner M, Erdmann E, Beuckelmann DJ. Calcium content of the sarcoplasmic reticulum in isolated ventricular myocytes from patients with terminal heart failure. J Mol Cell Cardiol. 1998;30:743–749. - PubMed
-
- Hobai IA, O’Rourke B. Decreased sarcoplasmic reticulum calcium content is responsible for defective excitation-contraction coupling in canine heart failure. Circulation. 2001;103:1577–1584. - PubMed
-
- Hasenfuss G, Pieske B. Calcium cycling in congestive heart failure. J Mol Cell Cardiol. 2002;34:951–969. - PubMed
-
- Houser SR, Margulies KB. Is depressed myocyte contractility centrally involved in heart failure? Circ Res. 2003;92:350–358. - PubMed
-
- Sipido KR, Eisner D. Something old, something new: changing views on the cellular mechanisms of heart failure. Cardiovasc Res. 2005;68:167–174. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
