

Genetic control of variegated KIR gene expression: polymorphisms of the bi-directional KIR3DL1 promoter are associated with distinct frequencies of gene expression
- PMID: 19008943
- PMCID: PMC2575236
- DOI: 10.1371/journal.pgen.1000254
Genetic control of variegated KIR gene expression: polymorphisms of the bi-directional KIR3DL1 promoter are associated with distinct frequencies of gene expression
Abstract
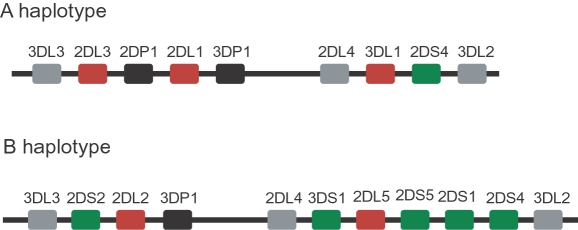
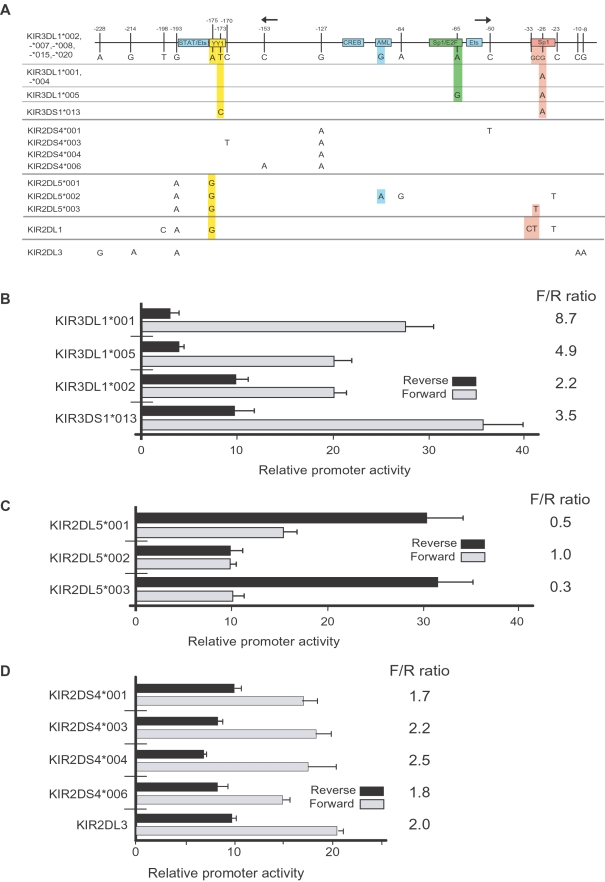
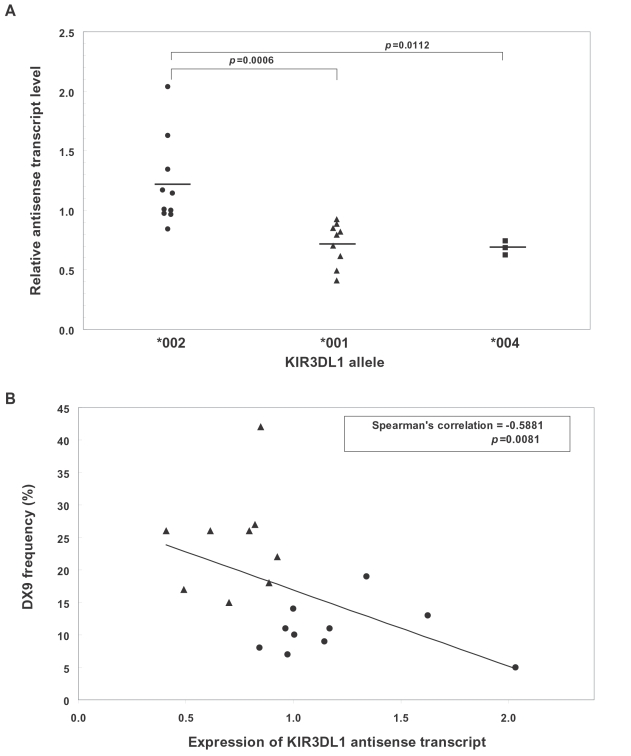
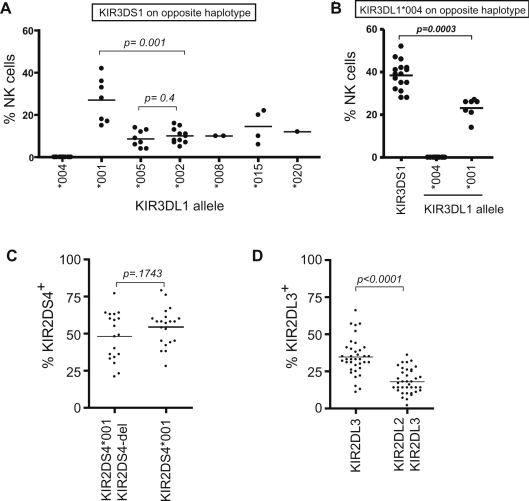
Natural killer (NK) cells play an important role in the detection and elimination of tumors and virus-infected cells by the innate immune system. Human NK cells use cell surface receptors (KIR) for class I MHC to sense alterations of class I on potential target cells. Individual NK cells only express a subset of the available KIR genes, generating specialized NK cells that can specifically detect alteration of a particular class I molecule or group of molecules. The probabilistic behavior of human KIR bi-directional promoters is proposed to control the frequency of expression of these variegated genes. Analysis of a panel of donors has revealed the presence of several functionally relevant promoter polymorphisms clustered mainly in the inhibitory KIR family members, especially the KIR3DL1 alleles. We demonstrate for the first time that promoter polymorphisms affecting the strength of competing sense and antisense promoters largely explain the differential frequency of expression of KIR3DL1 allotypes on NK cells. KIR3DL1/S1 subtypes have distinct biological activity and coding region variants of the KIR3DL1/S1 gene strongly influence pathogenesis of HIV/AIDS and other human diseases. We propose that the polymorphisms shown in this study to regulate the frequency of KIR3DL1/S1 subtype expression on NK cells contribute substantially to the phenotypic variation across allotypes with respect to disease resistance.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
-
- Lanier LL. NK cell recognition. Annu Rev Immunol. 2005;23:225–274. - PubMed
-
- Moretta L, Moretta A. Killer immunoglobulin-like receptors. Curr Opin Immunol. 2004;16:626–633. - PubMed
-
- Parham P. MHC class I molecules and KIRs in human history, health and survival. Nat Rev Immunol. 2005;5:201–214. - PubMed
-
- Yokoyama WM, Plougastel BF. Immune functions encoded by the natural killer gene complex. Nat Rev Immunol. 2003;3:304–316. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
