

Mesothelin-induced pancreatic cancer cell proliferation involves alteration of cyclin E via activation of signal transducer and activator of transcription protein 3
- PMID: 19010822
- PMCID: PMC2929833
- DOI: 10.1158/1541-7786.MCR-08-0095
Mesothelin-induced pancreatic cancer cell proliferation involves alteration of cyclin E via activation of signal transducer and activator of transcription protein 3
Abstract
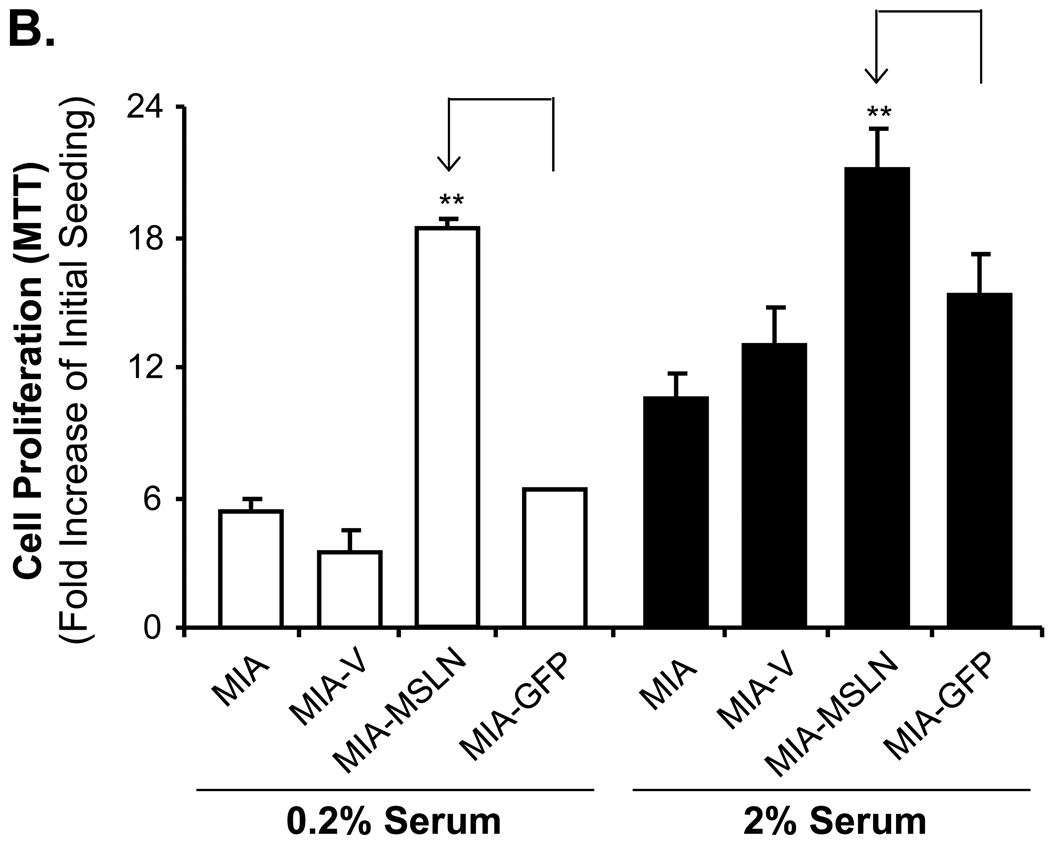
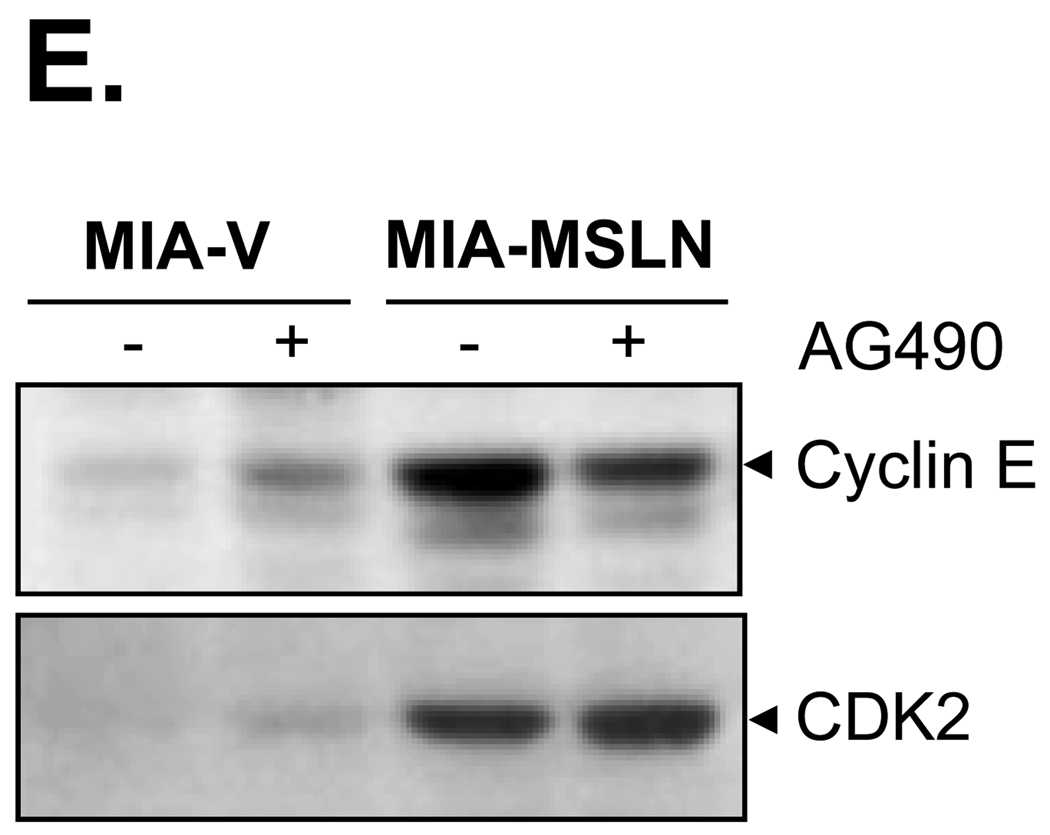
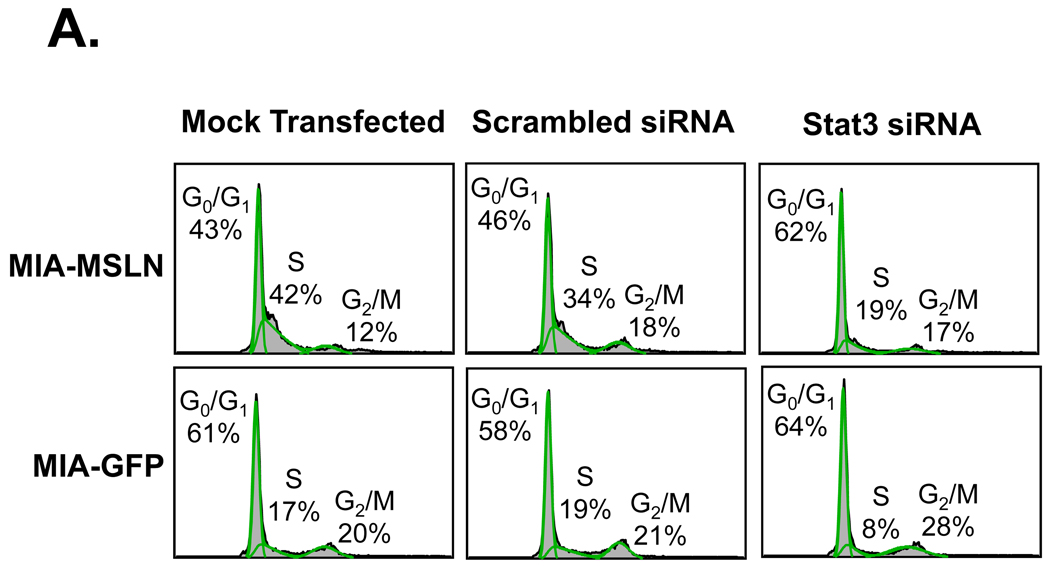
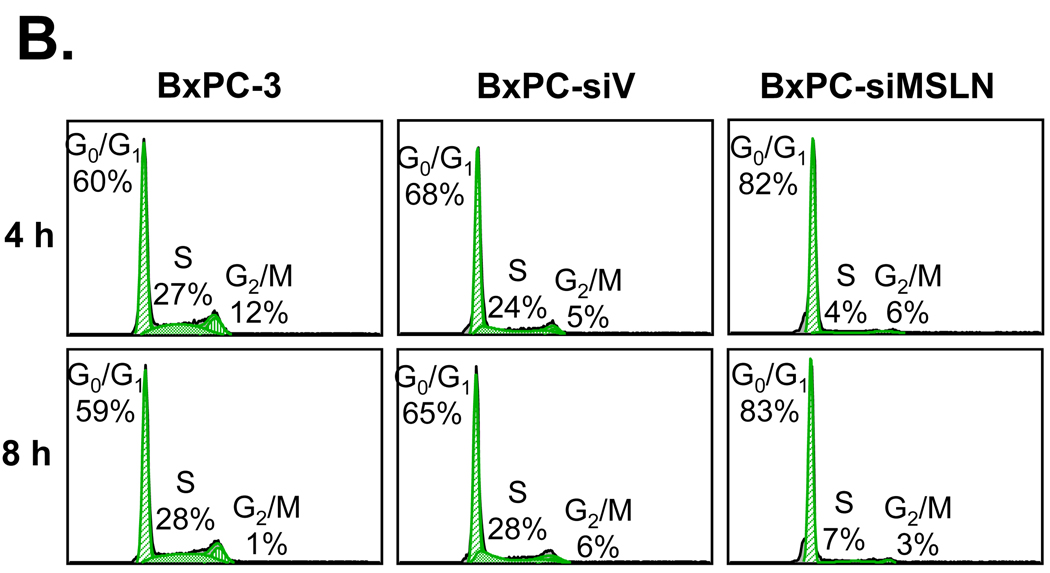
Mesothelin (MSLN) is a cell surface glycoprotein that is overexpressed in human pancreatic cancer. Although its value as a tumor marker for diagnosis and prognosis and as a preferred target of immunointervention has been evaluated, there is little information on the growth advantage of MSLN on tumor cells. In this study, we examined the effect of MSLN on pancreatic cancer cell proliferation, cell cycle progression, expression of cell cycle regulatory proteins, and signal transduction pathways in two pancreatic cancer cell lines, MIA-MSLN (overexpressing MSLN in MIA PaCa-2 cells) and BxPC-siMSLN (silencing MSLN in BxPC-3 cells). Increased cyclin E and cyclin-dependent kinase 2 expression found in MIA-MSLN cells correlated with significantly increased cell proliferation and faster cell cycle progression compared with control cells. BxPC-siMSLN cells showed slower proliferation and slower entry into the S phase than control cells. Signal transducer and activator of transcription protein 3 (Stat3) was constitutively activated in MIA-MSLN cells, but not in control cells. Inhibition of Stat3 activation in MIA-MSLN cells by the Janus-activated kinase-selective inhibitor tyrphostin AG490 was followed by a marked decrease in proliferation of the cells. Small interfering RNA against Stat3 significantly reduced the MIA-MSLN cell cycle progression with a concomitant decrease in cyclin E expression. Our data indicate that overexpression of MSLN in pancreatic cancer cells leads to constitutive activation of the transcription factor Stat3, which results in enhanced expression of cyclin E and cyclin E/cyclin-dependent kinase 2 complex formation as well as increased G(1)-S transition.
Conflict of interest statement
The authors do not have potential conflicts of interest.
Figures














References
-
- Hassan R, Bera T, Pastan I. Mesothelin: a new target for immunotherapy. Clin Cancer Res. 2004;10:3937–3942. - PubMed
-
- Chang K, Pai LH, Batra JK, Pastan I, Willingham MC. Characterization of the antigen (CAK1) recognized by monoclonal antibody K1 present on ovarian cancers and normal mesothelium. Cancer Res. 1992;52:181–186. - PubMed
-
- Argani P, Iacobuzio-Donahue C, Ryu B, et al. Mesothelin is overexpressed in the vast majority of ductal adenocarcinomas of the pancreas: identification of a new pancreatic cancer marker by serial analysis of gene expression (SAGE) Clin Cancer Res. 2001;7:3862–3868. - PubMed
-
- Ho M, Bera TK, Willingham MC, et al. Mesothelin expression in human lung cancer. Clin Cancer Res. 2007;13:1571–1575. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
