

Hydraulic failure defines the recovery and point of death in water-stressed conifers
- PMID: 19011001
- PMCID: PMC2613726
- DOI: 10.1104/pp.108.129783
Hydraulic failure defines the recovery and point of death in water-stressed conifers
Abstract
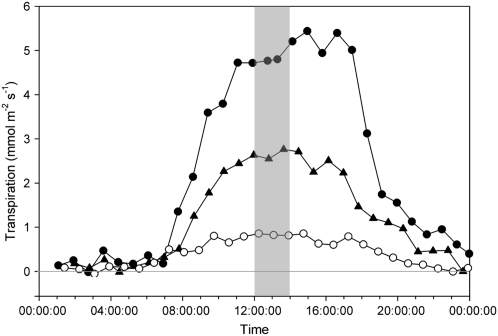
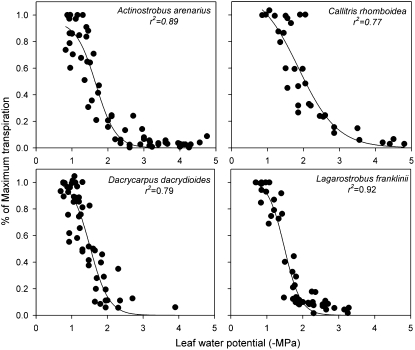
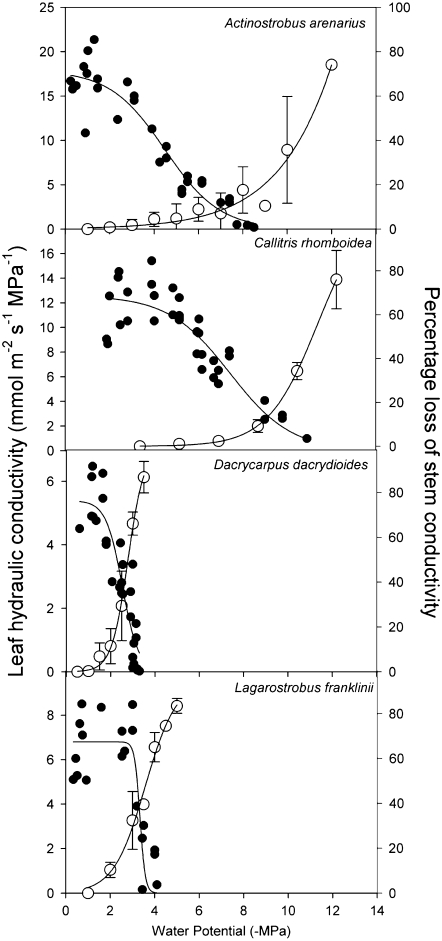
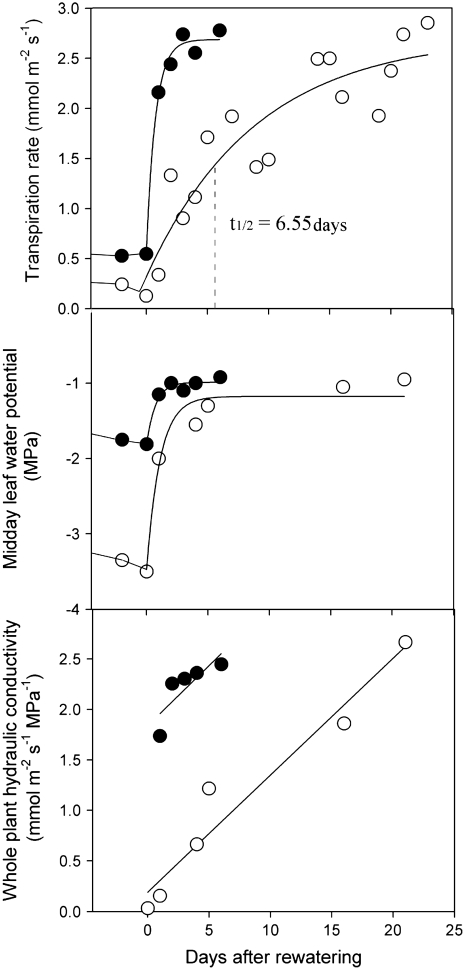
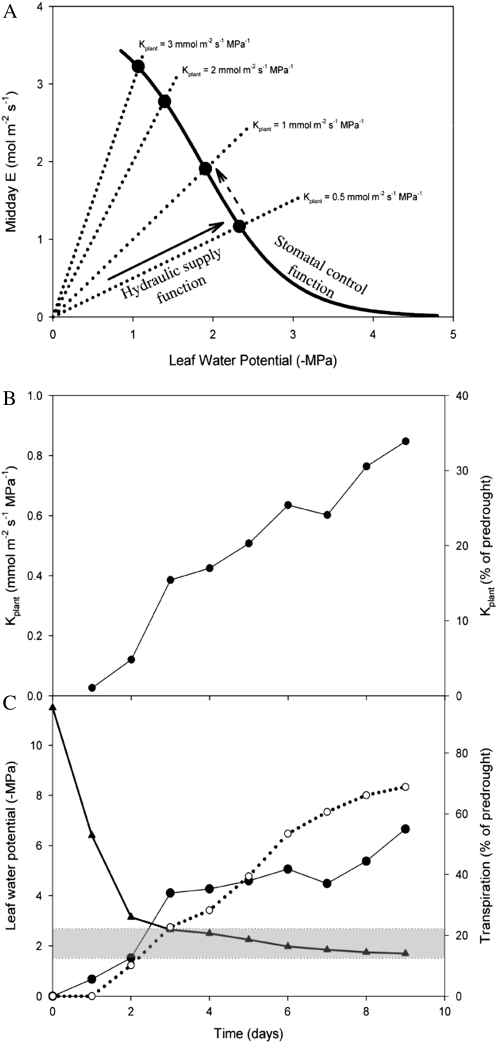
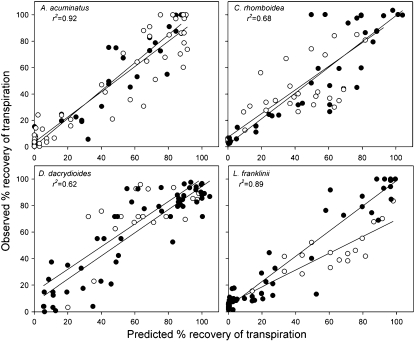
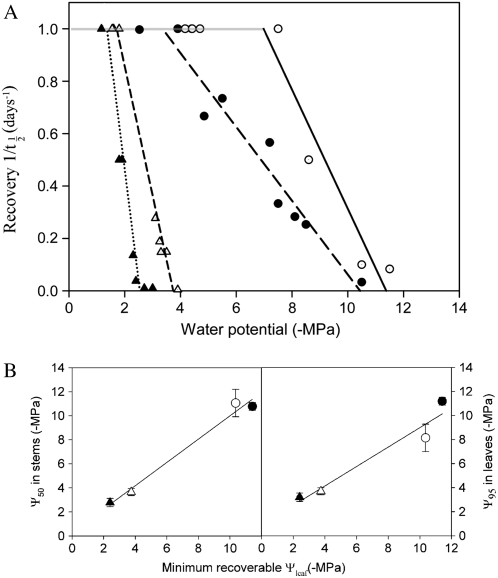
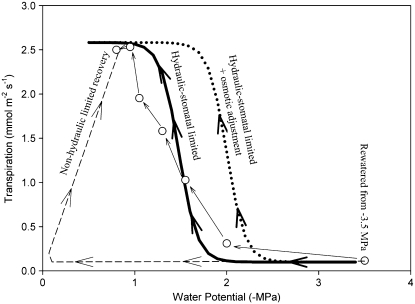
This study combines existing hydraulic principles with recently developed methods for probing leaf hydraulic function to determine whether xylem physiology can explain the dynamic response of gas exchange both during drought and in the recovery phase after rewatering. Four conifer species from wet and dry forests were exposed to a range of water stresses by withholding water and then rewatering to observe the recovery process. During both phases midday transpiration and leaf water potential (Psileaf) were monitored. Stomatal responses to Psileaf were established for each species and these relationships used to evaluate whether the recovery of gas exchange after drought was limited by postembolism hydraulic repair in leaves. Furthermore, the timing of gas-exchange recovery was used to determine the maximum survivable water stress for each species and this index compared with data for both leaf and stem vulnerability to water-stress-induced dysfunction measured for each species. Recovery of gas exchange after water stress took between 1 and >100 d and during this period all species showed strong 1:1 conformity to a combined hydraulic-stomatal limitation model (r2 = 0.70 across all plants). Gas-exchange recovery time showed two distinct phases, a rapid overnight recovery in plants stressed to <50% loss of leaf hydraulic conductance (Kleaf) and a highly Psileaf-dependent phase in plants stressed to >50% loss of Kleaf. Maximum recoverable water stress (Psimin) corresponded to a 95% loss of Kleaf. Thus, we conclude that xylem hydraulics represents a direct limit to the drought tolerance of these conifer species.
Figures
References
-
- Alder NN, Pockman WT, Sperry JS, Nuismer S (1997) Use of centrifugal force in the study of xylem cavitation. J Exp Bot 48 665–674
-
- Breda N, Huc R, Granier A, Dreyer E (2006) Temperate forest trees and stands under severe drought: a review of ecophysiological responses, adaptation processes and long-term consequences. Ann For Sci 63 625–644
-
- Brodribb TJ, Feild TS (2000) Stem hydraulic supply is linked to leaf photosynthetic capacity: evidence from New Caledonian and Tasmanian rainforests. Plant Cell Environ 23 1381–1388
-
- Brodribb TJ, Hill RS (1999) The importance of xylem constraints in the distribution of conifer species. New Phytol 143 365–372
-
- Brodribb TJ, Holbrook NM (2003) Changes in leaf hydraulic conductance during leaf shedding in seasonally dry tropical forest. New Phytol 158 295–303
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
