

Motor modulation of afferent somatosensory circuits
- PMID: 19011625
- PMCID: PMC2597103
- DOI: 10.1038/nn.2227
Motor modulation of afferent somatosensory circuits
Abstract
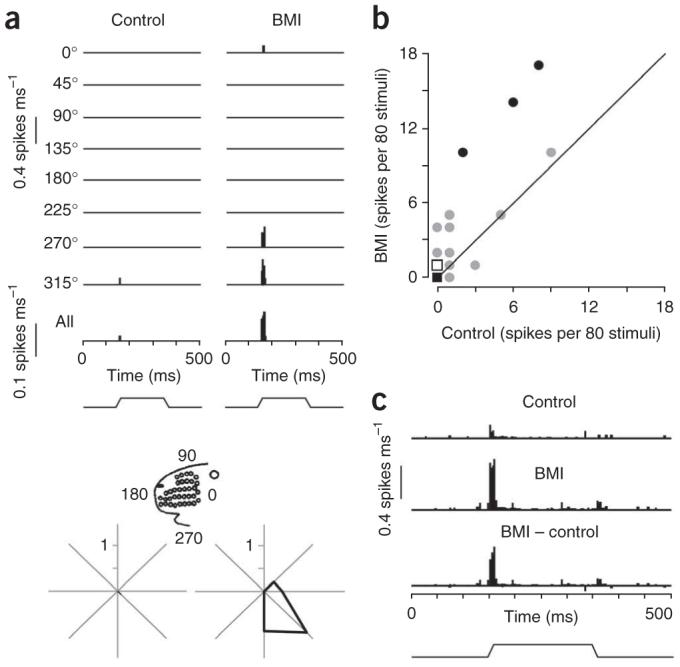
A prominent feature of thalamocortical circuitry in sensory systems is the extensive and highly organized feedback projection from the cortex to the thalamic neurons that provide stimulus-specific input to the cortex. In lightly sedated rats, we found that focal enhancement of motor cortex activity facilitated sensory-evoked responses of topographically aligned neurons in primary somatosensory cortex, including antidromically identified corticothalamic cells; similar effects were observed in ventral posterior medial thalamus (VPm). In behaving rats, thalamic responses were normally smaller during whisking but larger when signal transmission in brainstem trigeminal nuclei was bypassed or altered. During voluntary movement, sensory activity may be globally suppressed in the brainstem, whereas signaling by cortically facilitated VPm neurons is simultaneously enhanced relative to other VPm neurons receiving no such facilitation.
Figures







Comment in
-
And motion changes it all.Nat Neurosci. 2008 Dec;11(12):1369-70. doi: 10.1038/nn1208-1369. Nat Neurosci. 2008. PMID: 19023341 No abstract available.
References
-
- Ahissar E, Kleinfeld D. Closed-loop neuronal computations: focus on vibrissa somatosensation in rat. Cereb. Cortex. 2003;13:53–62. - PubMed
-
- Carvell GE, Miller SA, Simons DJ. The relationship of vibrissal motor cortex unit activity to whisking in the awake rat. Somatosens. Mot. Res. 1996;13:115–127. - PubMed
-
- Krupa DJ, Wiest MC, Shuler MG, Laubach M, Nicolelis MA. Layer-specific somatosensory cortical activation during active tactile discrimination. Science. 2004;304:1989–1992. - PubMed
-
- Hentschke H, Haiss F, Schwarz C. Central signals rapidly switch tactile processing in rat barrel cortex during whisker movements. Cereb. Cortex. 2006;16:1142–1156. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
