

An oncogenomics-based in vivo RNAi screen identifies tumor suppressors in liver cancer
- PMID: 19012953
- PMCID: PMC2990916
- DOI: 10.1016/j.cell.2008.09.061
An oncogenomics-based in vivo RNAi screen identifies tumor suppressors in liver cancer
Abstract
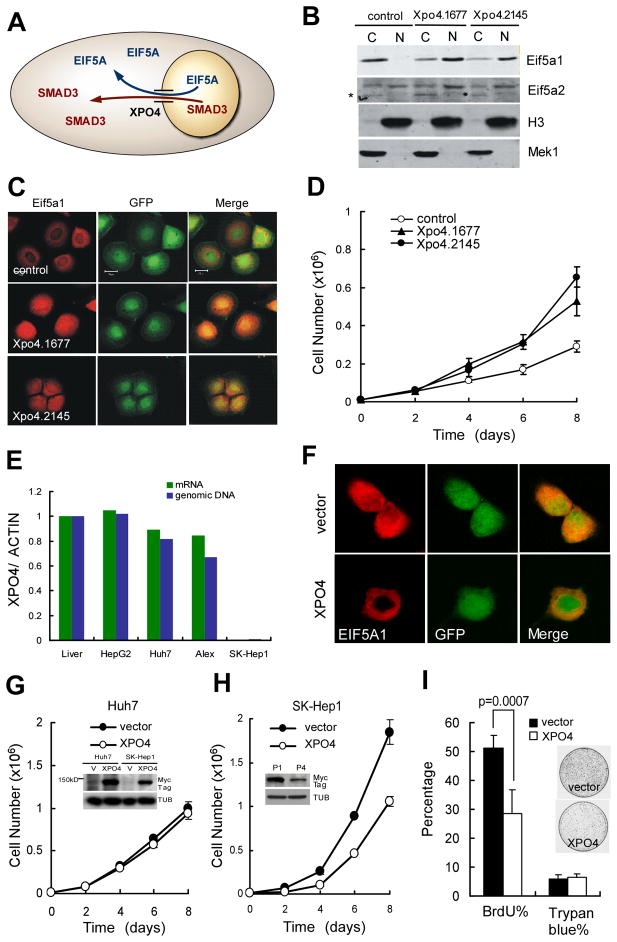
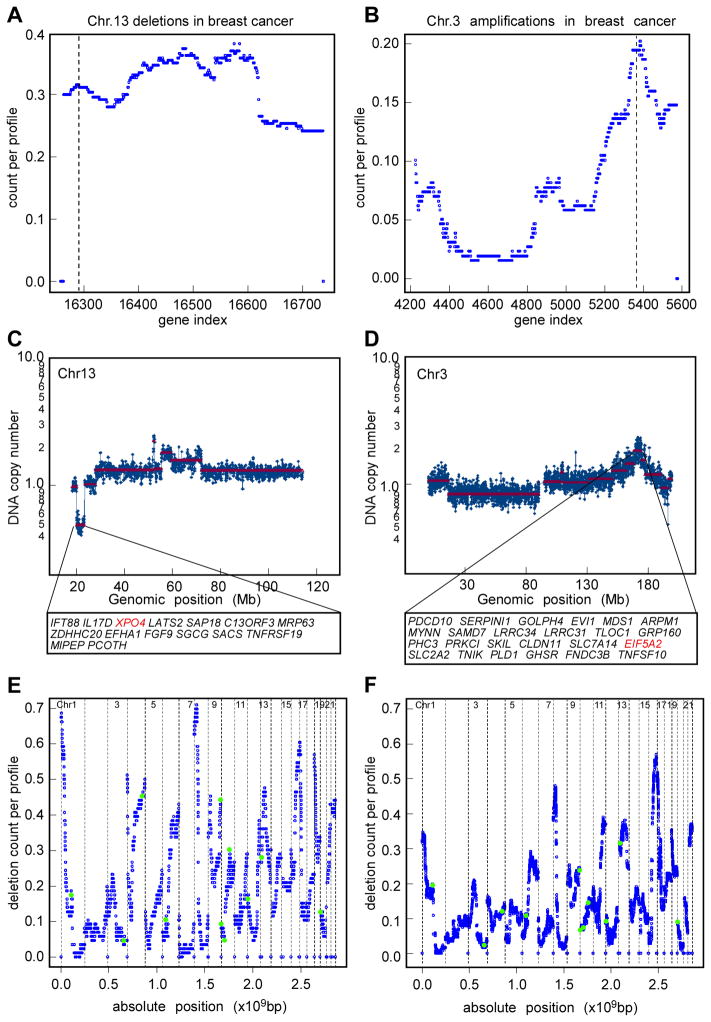
Cancers are highly heterogeneous and contain many passenger and driver mutations. To functionally identify tumor suppressor genes relevant to human cancer, we compiled pools of short hairpin RNAs (shRNAs) targeting the mouse orthologs of genes recurrently deleted in a series of human hepatocellular carcinomas and tested their ability to promote tumorigenesis in a mosaic mouse model. In contrast to randomly selected shRNA pools, many deletion-specific pools accelerated hepatocarcinogenesis in mice. Through further analysis, we identified and validated 13 tumor suppressor genes, 12 of which had not been linked to cancer before. One gene, XPO4, encodes a nuclear export protein whose substrate, EIF5A2, is amplified in human tumors, is required for proliferation of XPO4-deficient tumor cells, and promotes hepatocellular carcinoma in mice. Our results establish the feasibility of in vivo RNAi screens and illustrate how combining cancer genomics, RNA interference, and mosaic mouse models can facilitate the functional annotation of the cancer genome.
Figures





Comment in
-
RNAi Delivers Insights into Liver Cancer.Cell. 2008 Nov 28;135(5):793-5. doi: 10.1016/j.cell.2008.11.001. Cell. 2008. PMID: 19041743
References
-
- Benne R, Hershey JW. The mechanism of action of protein synthesis initiation factors from rabbit reticulocytes. J Biol Chem. 1978;253:3078–3087. - PubMed
-
- Caraglia M, Marra M, Giuberti G, D'Alessandro AM, Budillon A, Del Prete S, Lentini A, Beninati S, Abbruzzese A. The role of eukaryotic initiation factor 5A in the control of cell proliferation and apoptosis. Amino Acids. 2001;20:91–104. - PubMed
-
- Chow LM, Baker SJ. PTEN function in normal and neoplastic growth. Cancer Lett. 2006;241:184–196. - PubMed
-
- Clement PM, Henderson CA, Jenkins ZA, Smit-McBride Z, Wolff EC, Hershey JW, Park MH, Johansson HE. Identification and characterization of eukaryotic initiation factor 5A-2. Eur J Biochem. 2003;270:4254–4263. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
