

Low-frequency neuronal oscillations as instruments of sensory selection
- PMID: 19012975
- PMCID: PMC2990947
- DOI: 10.1016/j.tins.2008.09.012
Low-frequency neuronal oscillations as instruments of sensory selection
Abstract
Neuroelectric oscillations reflect rhythmic shifting of neuronal ensembles between high and low excitability states. In natural settings, important stimuli often occur in rhythmic streams, and when oscillations entrain to an input rhythm their high excitability phases coincide with events in the stream, effectively amplifying neuronal input responses. When operating in a 'rhythmic mode', attention can use these differential excitability states as a mechanism of selection by simply enforcing oscillatory entrainment to a task-relevant input stream. When there is no low-frequency rhythm that oscillations can entrain to, attention operates in a 'continuous mode', characterized by extended increase in gamma synchrony. We review the evidence for early sensory selection by oscillatory phase-amplitude modulations, its mechanisms and its perceptual and behavioral consequences.
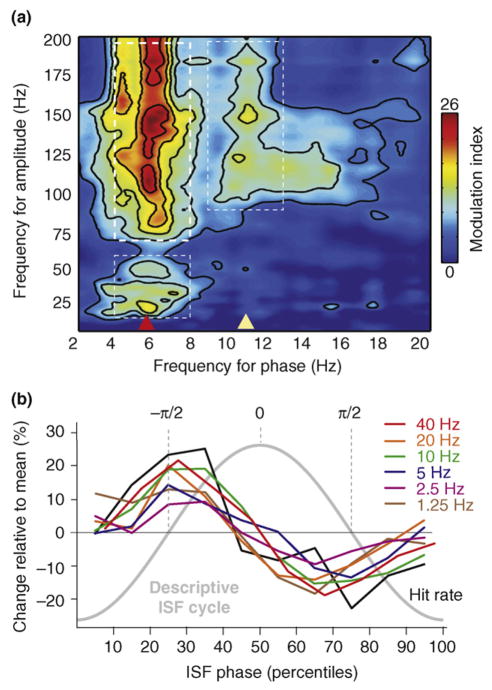
Figures
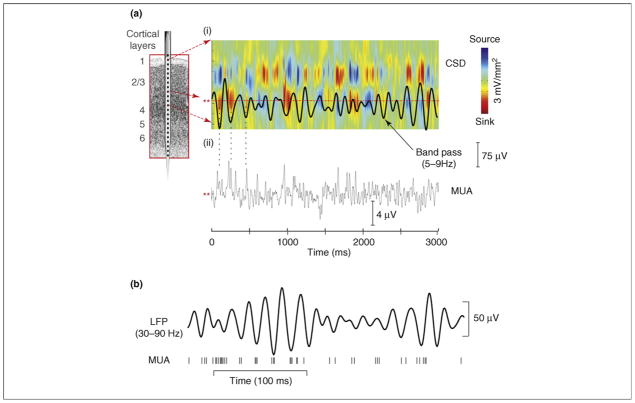
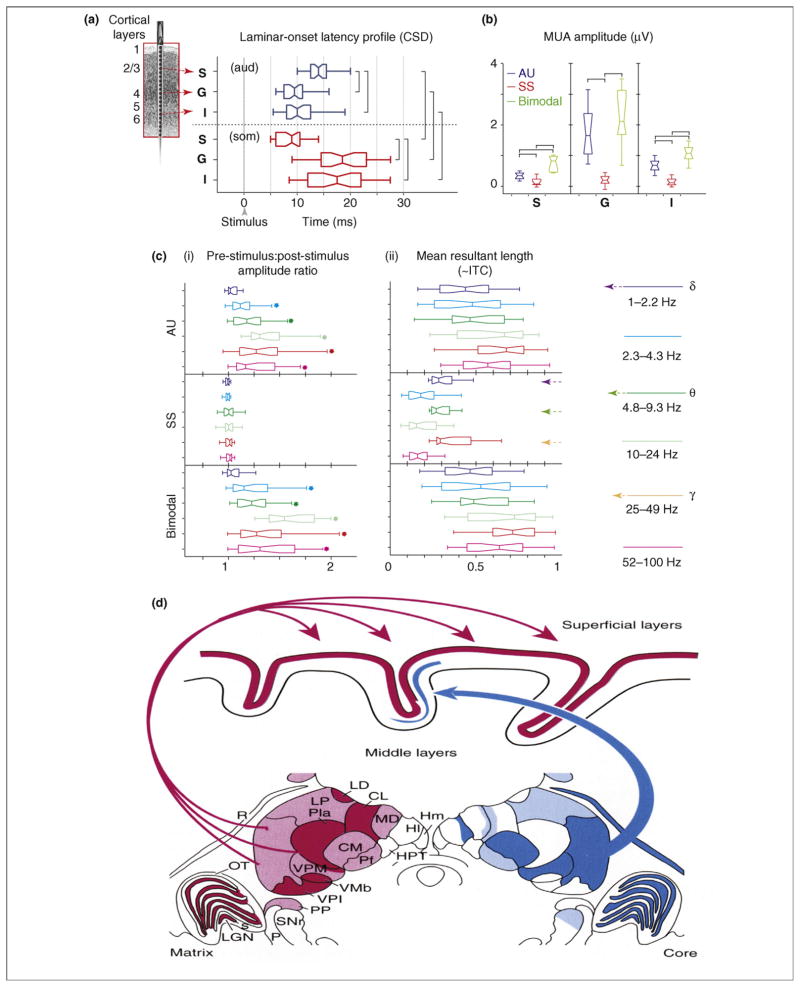
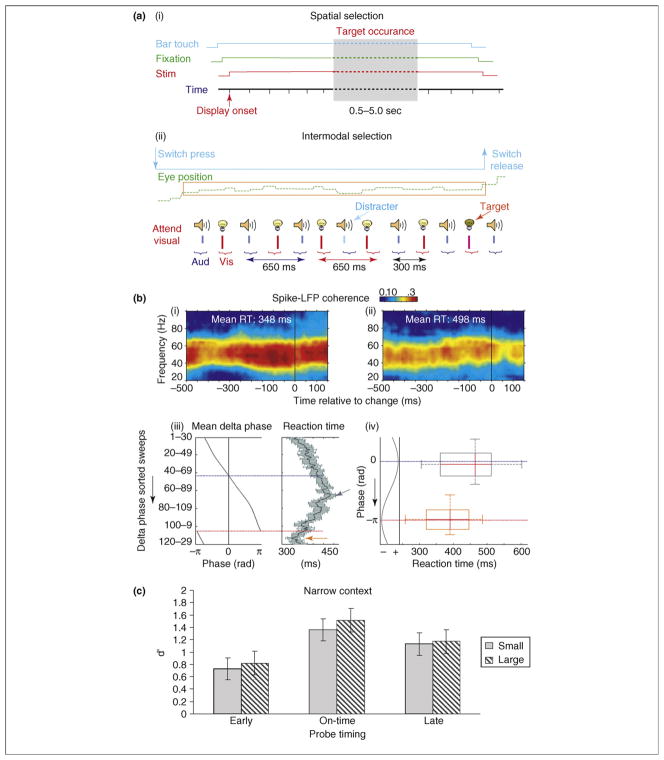
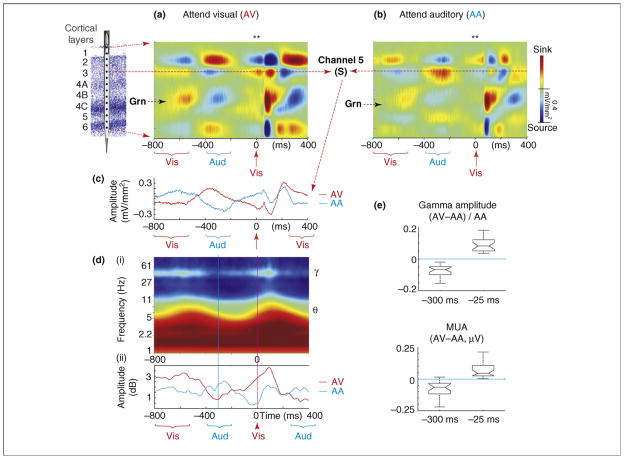
References
-
- Bishop G. Cyclical changes in excitability of the optic pathway of the rabbit. Am J Physiol. 1933;103:213–224.
-
- Maurer AP, McNaughten BL. Network and intrinsic cellular mechanisms underlying theta phase precession of hippocampal neurons. Trends Neurosci. 2007;30:325–333. - PubMed
-
- Jensen O, et al. Human gamma-frequency oscillations associated with attention and memory. Trends Neurosci. 2007;30:317–324. - PubMed
-
- Palva S, Palva JM. New vistas for α frequency band oscillations. Trends Neurosci. 2007;30:150–158. - PubMed
-
- Singer W, Gray CM. Visual feature integration and the temporal correlation hypothesis. Annu Rev Neurosci. 1995;18:555–586. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
