

Cockroaches keep predators guessing by using preferred escape trajectories
- PMID: 19013065
- PMCID: PMC2678410
- DOI: 10.1016/j.cub.2008.09.062
Cockroaches keep predators guessing by using preferred escape trajectories
Abstract
Antipredator behavior is vital for most animals and calls for accurate timing and swift motion. Whereas fast reaction times [1] and predictable, context-dependent escape-initiation distances [2] are common features of most escape systems, previous work has highlighted the need for unpredictability in escape directions, in order to prevent predators from learning a repeated, fixed pattern [3-5]. Ultimate unpredictability would result from random escape trajectories. Although this strategy would deny any predictive power to the predator, it would also result in some escape trajectories toward the threat. Previous work has shown that escape trajectories are in fact generally directed away from the threat, although with a high variability [5-8]. However, the rules governing this variability are largely unknown. Here, we demonstrate that individual cockroaches (Periplaneta americana, a much-studied model prey species [9-14]) keep each escape unpredictable by running along one of a set of preferred trajectories at fixed angles from the direction of the threatening stimulus. These results provide a new paradigm for understanding the behavioral strategies for escape responses, underscoring the need to revisit the neural mechanisms controlling escape directions in the cockroach and similar animal models, and the evolutionary forces driving unpredictable, or "protean"[3], antipredator behavior.
Figures
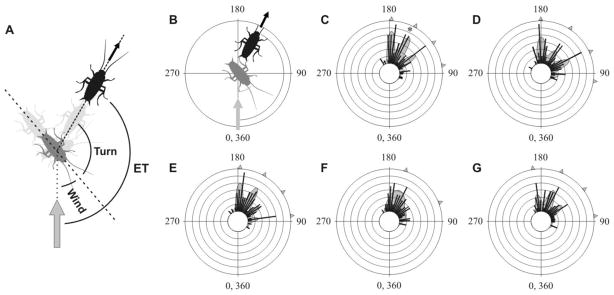
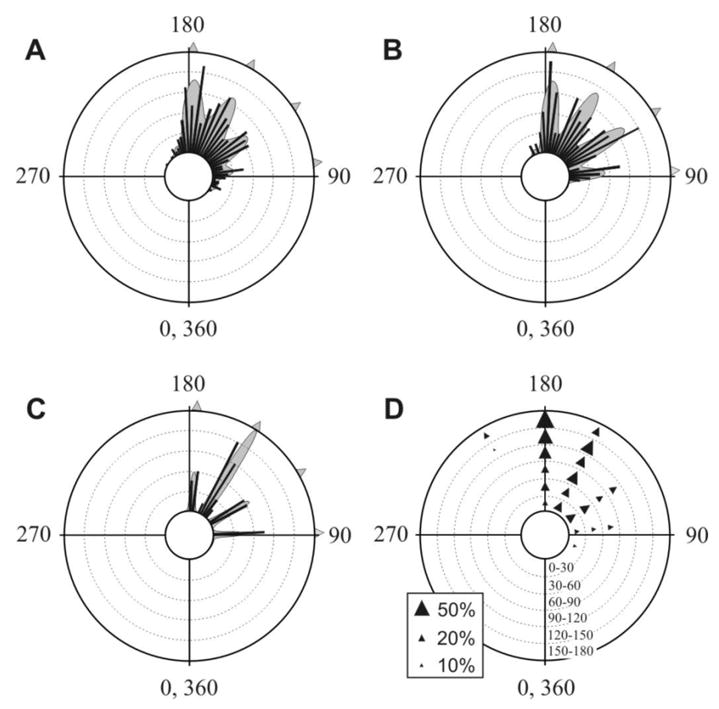
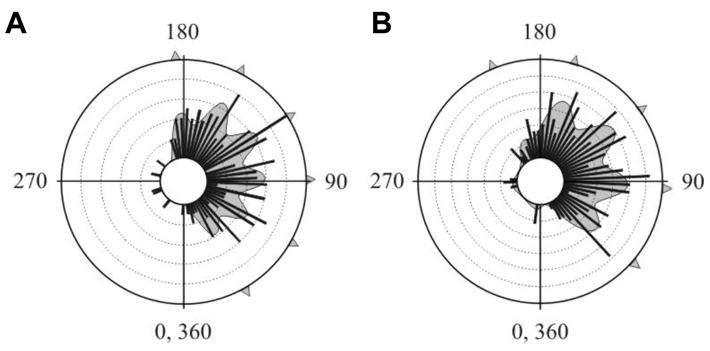
Comment in
-
Behavioral biology: inside the mind of proteus?Curr Biol. 2009 Jan 13;19(1):R27-8. doi: 10.1016/j.cub.2008.11.010. Curr Biol. 2009. PMID: 19138587 Review.
References
-
- Bullock TH. Comparative neuroethology of startle, rapid escape and giant fiber-mediated responses. In: Eaton RC, editor. Neural Mechanisms of Startle Behaviour. New York: Plenum Press; 1984. pp. 1–13.
-
- Ydenberg RC, Dill LM. The economics of fleeing from predators. Advances in the Study of Behaviour. 1986;16:229–249.
-
- Humphries DA, Driver PM. Protean defence by prey animals. Oecologia. 1970;5:285–302. - PubMed
-
- Driver PM, Humphries DA. Protean Behaviour: The Biology of Unpredictability. Oxford: Clarendon; 1988.
-
- Godin J-GJ. Evading predators. In: Godin J-GJ, editor. Behavioural Ecology of Teleost Fishes. Oxford: Oxford University Press; 1997. pp. 191–236.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
