

MicroRNA processing pathway regulates olfactory neuron morphogenesis
- PMID: 19013069
- PMCID: PMC2612040
- DOI: 10.1016/j.cub.2008.09.045
MicroRNA processing pathway regulates olfactory neuron morphogenesis
Abstract
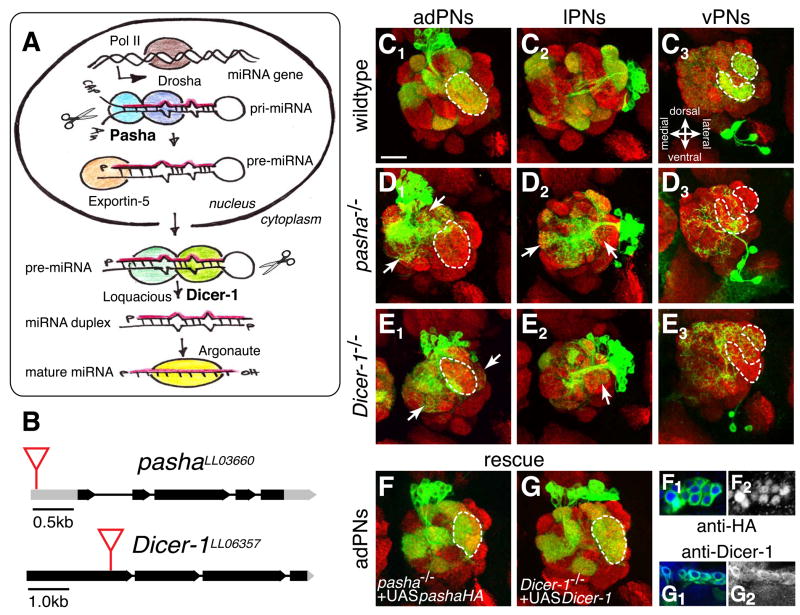
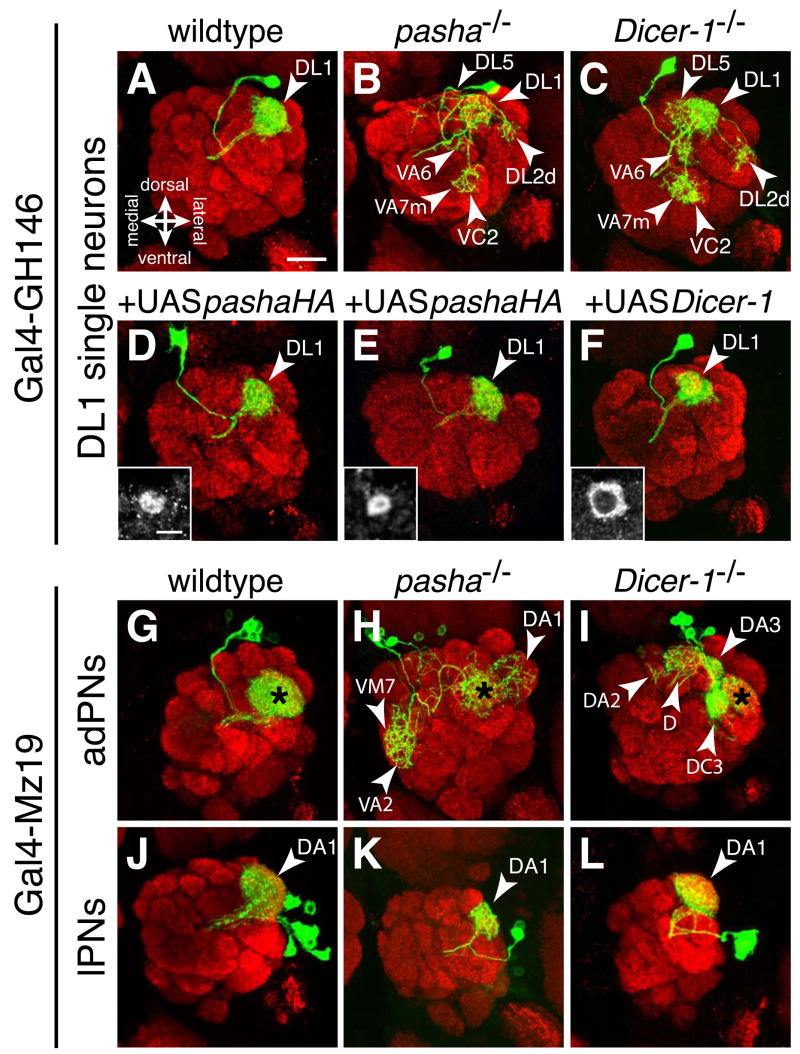
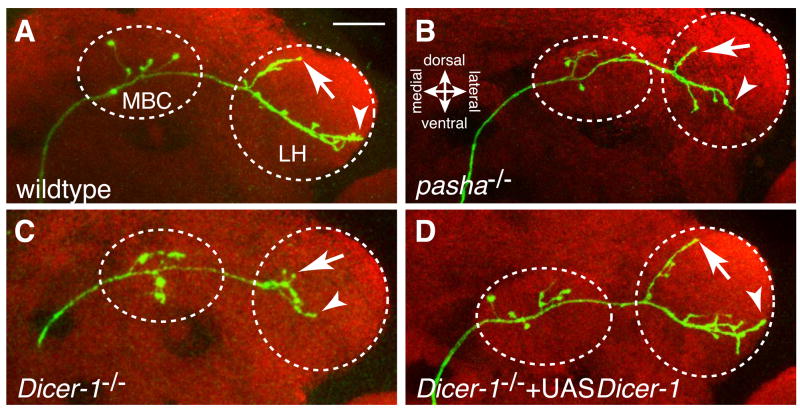
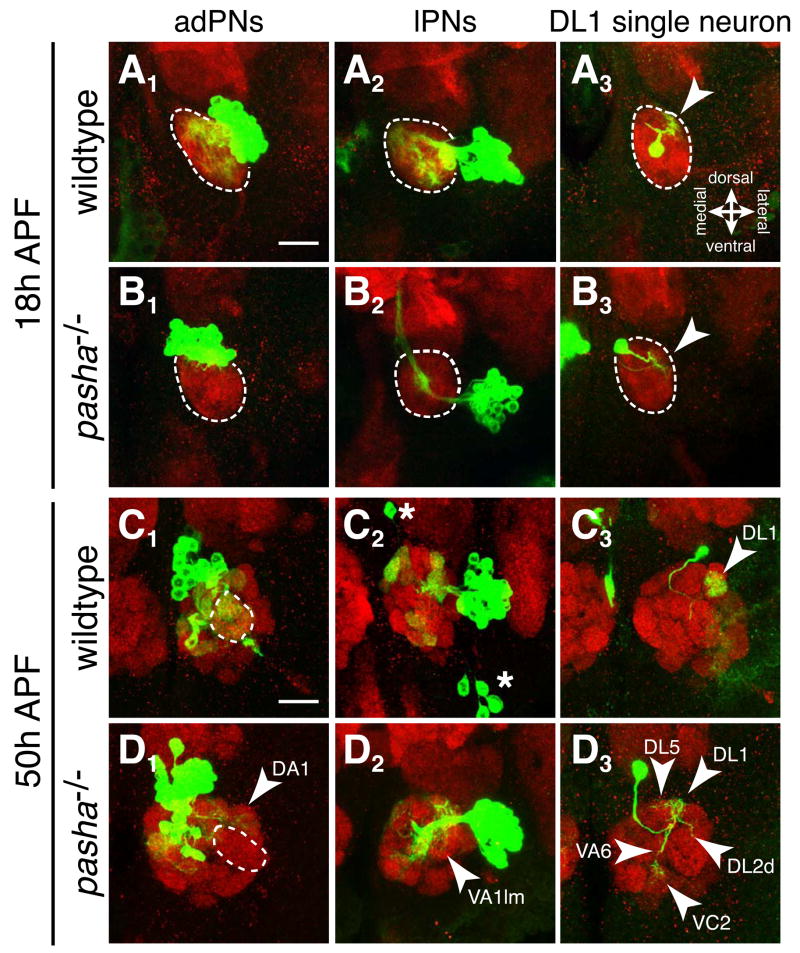
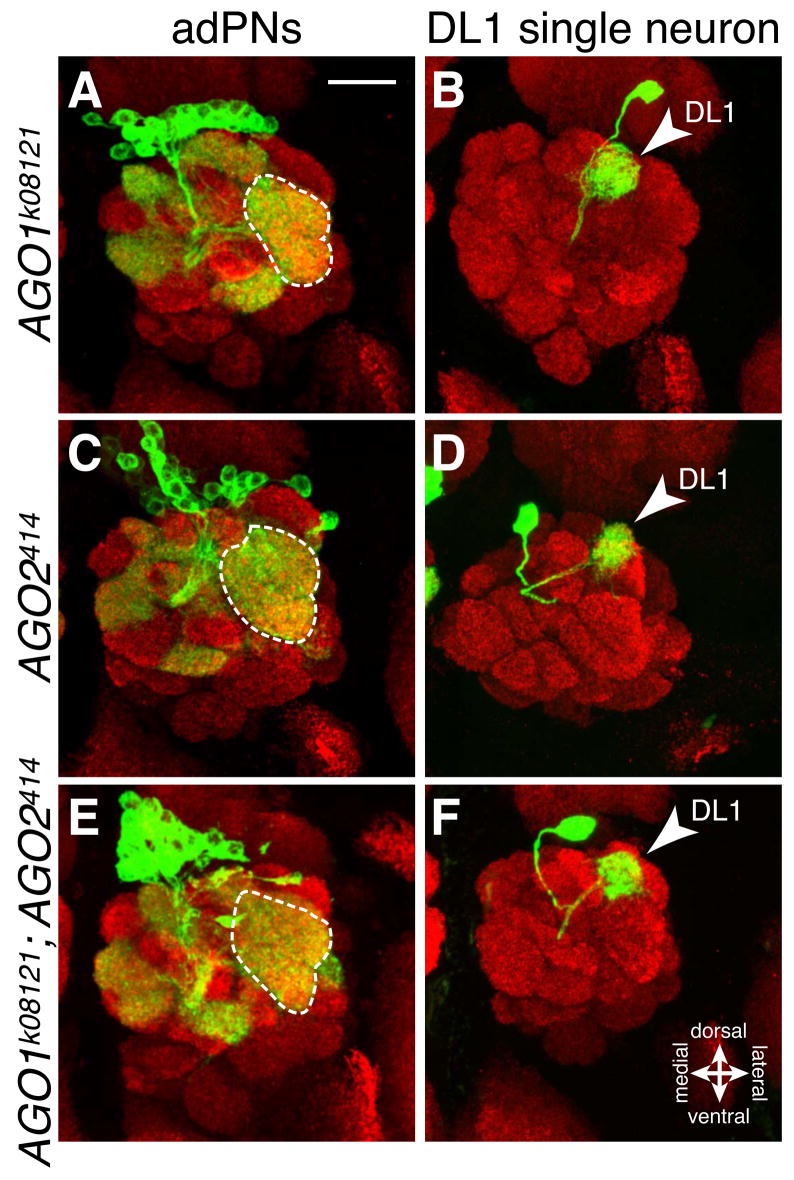
The microRNA (miRNA) processing pathway produces miRNAs as posttranscriptional regulators of gene expression. The nuclear RNase III Drosha catalyzes the first processing step together with the dsRNA binding protein DGCR8/Pasha generating pre-miRNAs [1, 2]. The next cleavage employs the cytoplasmic RNase III Dicer producing miRNA duplexes [3, 4]. Finally, Argonautes are recruited with miRNAs into an RNA-induced silencing complex for mRNA recognition (Figure 1A). Here, we identify two members of the miRNA pathway, Pasha and Dicer-1, in a forward genetic screen for mutations that disrupt wiring specificity of Drosophila olfactory projection neurons (PNs). The olfactory system is built as discrete map of highly stereotyped neuronal connections [5, 6]. Each PN targets dendrites to a specific glomerulus in the antennal lobe and projects axons stereotypically into higher brain centers [7-9]. In selected PN classes, pasha and Dicer-1 mutants cause specific PN dendrite mistargeting in the antennal lobe and altered axonal terminations in higher brain centers. Furthermore, Pasha and Dicer-1 act cell autonomously in postmitotic neurons to regulate dendrite and axon targeting during development. However, Argonaute-1 and Argonaute-2 are dispensable for PN morphogenesis. Our findings suggest a role for the miRNA processing pathway in establishing wiring specificity in the nervous system.
Figures
References
-
- Denli AM, Tops BB, Plasterk RH, Ketting RF, Hannon GJ. Processing of primary microRNAs by the Microprocessor complex. Nature. 2004;432:231–235. - PubMed
-
- Gregory RI, Yan KP, Amuthan G, Chendrimada T, Doratotaj B, Cooch N, Shiekhattar R. The Microprocessor complex mediates the genesis of microRNAs. Nature. 2004;432:235–240. - PubMed
-
- Hutvagner G, McLachlan J, Pasquinelli AE, Balint E, Tuschl T, Zamore PD. A cellular function for the RNA-interference enzyme Dicer in the maturation of the let-7 small temporal RNA. Science. 2001;293:834–838. - PubMed
-
- Axel R. The molecular logic of smell. Sci Am. 1995;273:154–159. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
