

Evidence for activation of the TGF-beta1 promoter by C/EBPbeta and its modulation by Smads
- PMID: 19014341
- PMCID: PMC2988467
- DOI: 10.1089/jir.2008.0036
Evidence for activation of the TGF-beta1 promoter by C/EBPbeta and its modulation by Smads
Abstract
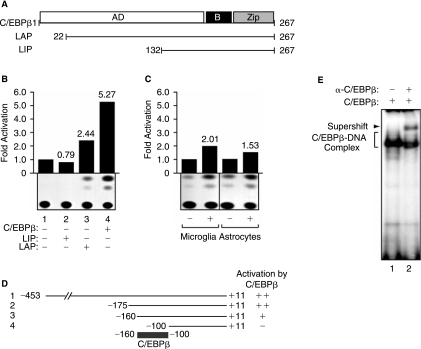
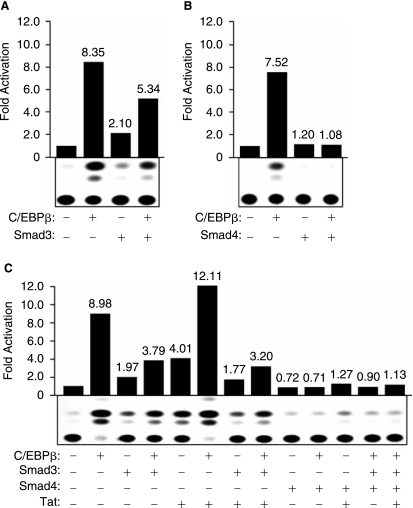
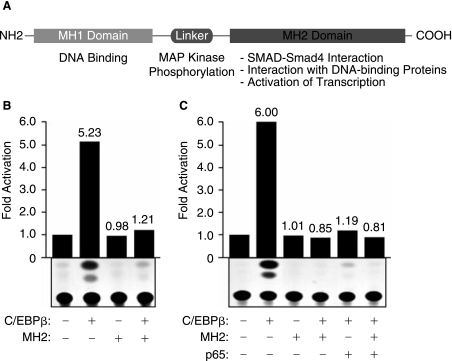
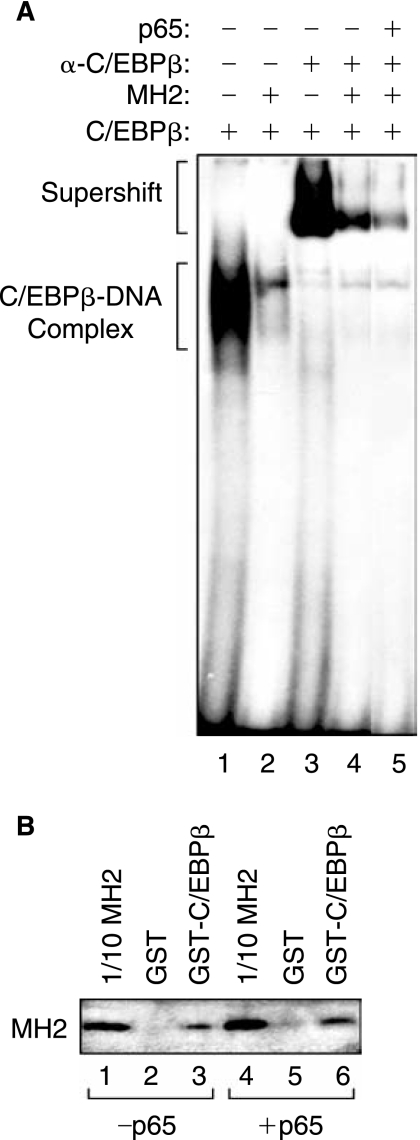
The transforming growth factor-beta1 (TGF-beta1) is a cytokine involved in many biological events inlcuding immunosuppression, angiogenesis, cell growth, and apoptosis. Expression of TGF-beta1 at the transcriptional level is controlled by a series of ubiquitous and specialized factors whose activities can be modulated by a variety of signaling events. Here we demonstrate that activity of the TGF-beta1 promoter is increased by C/EBPbeta, a DNA-binding transcription factor whose activity can be influenced by several immunomodulators, in astrocytes and microglial cells. Interestingly, expression of Smad3 and Smad4, the downstream regulators of the TGF-beta1-signaling pathway, impairs the activity of C/EBPbeta on the TGF-beta1 promoter. Further, we demonstrate that MH2, a common domain among Smads that has protein-binding activities, interacts with C/EBPbeta and decreases its association with a region of the TGF-beta1 promoter that is responsive to C/EBPbeta activation. Interestingly, the p65 subunit of nuclear factor-kappaB (NF-kappaB), which also interacts with C/EBPbeta, cooperates with MH2 and decreased DNA-binding and transcriptional activities of C/EBPbeta on the TGF-beta1 promoter. These observations indicate that an autoregulatory mechanism, involving the MH2 domain of Smads, modulates activation of the TGF-beta1 promoter by C/EBPbeta. Further, our results show that the interplay between NF-kappaB and C/EBPbeta has an impact on the ability of C/EBPbeta to stimulate TGF-beta1 transcription, hence, suggesting that the cross-communication of signaling pathways that modulate NF-kappaB and C/EBPbeta may dictate the level of TGF-beta1 promoter activity.
Figures
References
-
- Abraham S. Sweet T. Sawaya BE. Rappaport J. Khalili K. Amini S. Cooperative interaction of C/EBP beta and Tat modulates MCP-1 gene transcription in astrocytes. J Neuroimmunol. 2005;160:219–227. - PubMed
-
- Amini S. Clavo A. Nadraga Y. Giordano A. Khalili K. Sawaya BE. Interplay between cdk9 and NF-kappaB factors determines the level of HIV-1 gene transcription in astrocytic cells. Oncogene. 2002;21:5797–5803. - PubMed
-
- Coyle-Rink J. Sweet T. Abraham S. Sawaya B. Batuman O. Khalili K. Amini S. Interaction between TGFbeta signaling proteins and C/EBP controls basal and Tat-mediated transcription of HIV-1 LTR in astrocytes. Virology. 2002;299:240–247. - PubMed
-
- Descombes P. Schibler U. A liver-enriched transcriptional activator protein, LAP, and a transcriptional inhibitory protein, LIP, are translated from the same mRNA. Cell. 1991;67:569–579. - PubMed
-
- Eldeen MB. Deshmane SL. Simbiri K. Khalili K. Amini S. Sawaya BE. MH2 domain of Smad3 reduces HIV-1 Tat-induction of cytokine secretion. J Neuroimmunol. 2006;176:174–180. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
