

A novel expression platform for the production of diabetes-associated autoantigen human glutamic acid decarboxylase (hGAD65)
- PMID: 19014643
- PMCID: PMC2621204
- DOI: 10.1186/1472-6750-8-87
A novel expression platform for the production of diabetes-associated autoantigen human glutamic acid decarboxylase (hGAD65)
Abstract
Background: Human glutamic acid decarboxylase 65 (hGAD65) is a key autoantigen in type 1 diabetes, having much potential as an important marker for the prediction and diagnosis of type 1 diabetes, and for the development of novel antigen-specific therapies for the treatment of type 1 diabetes. However, recombinant production of hGAD65 using conventional bacterial or mammalian cell culture-based expression systems or nuclear transformed plants is limited by low yield and low efficiency. Chloroplast transformation of the unicellular eukaryotic alga Chlamydomonas reinhardtii may offer a potential solution.
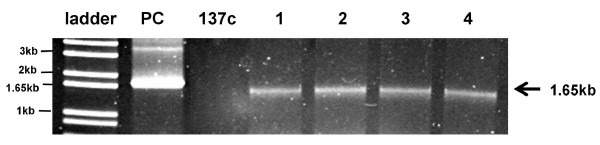
Results: A DNA cassette encoding full-length hGAD65, under the control of the C. reinhardtii chloroplast rbcL promoter and 5'- and 3'-UTRs, was constructed and introduced into the chloroplast genome of C. reinhardtii by particle bombardment. Integration of hGAD65 DNA into the algal chloroplast genome was confirmed by PCR. Transcriptional expression of hGAD65 was demonstrated by RT-PCR. Immunoblotting verified the expression and accumulation of the recombinant protein. The antigenicity of algal-derived hGAD65 was demonstrated with its immunoreactivity to diabetic sera by ELISA and by its ability to induce proliferation of spleen cells from NOD mice. Recombinant hGAD65 accumulated in transgenic algae, accounts for approximately 0.25-0.3% of its total soluble protein.
Conclusion: Our results demonstrate the potential value of C. reinhardtii chloroplasts as a novel platform for rapid mass production of immunologically active hGAD65. This demonstration opens the future possibility for using algal chloroplasts as novel bioreactors for the production of many other biologically active mammalian therapeutic proteins.
Figures






References
-
- Goddijn OJM, Pen J. Plants as bioreactors. Trends Biotechnol. 1995;13:379–387. doi: 10.1016/S0167-7799(00)88985-4. - DOI
-
- Ma SW, Tremblay R, Wang XF, Jevnikar AM. Plant-based pharmaceuticals and its application in oral tolerance. In: Pontell EB, editor. Immune Tolerance Research Development. New York: Nova Science Publishers; 2008. pp. 79–95.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
