

USP19 deubiquitinating enzyme supports cell proliferation by stabilizing KPC1, a ubiquitin ligase for p27Kip1
- PMID: 19015242
- PMCID: PMC2612522
- DOI: 10.1128/MCB.00329-08
USP19 deubiquitinating enzyme supports cell proliferation by stabilizing KPC1, a ubiquitin ligase for p27Kip1
Erratum in
- Mol Cell Biol. 2009 Jun;29(11):3241
Abstract
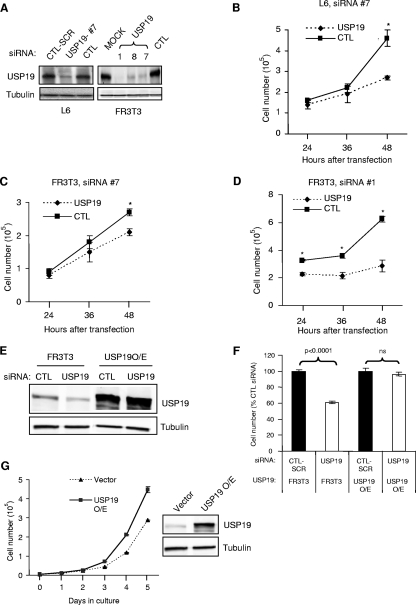
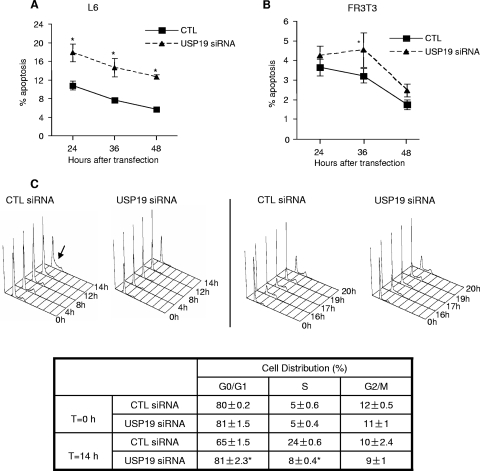
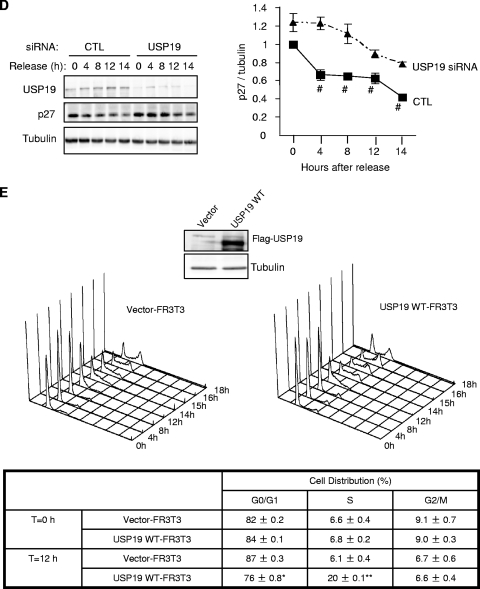
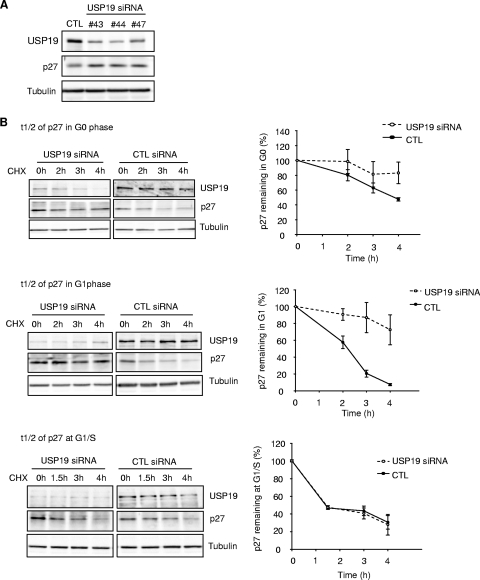
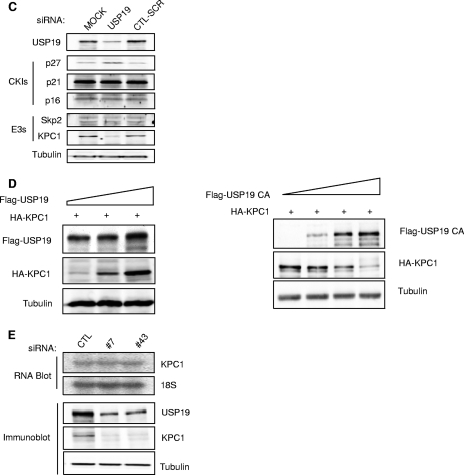
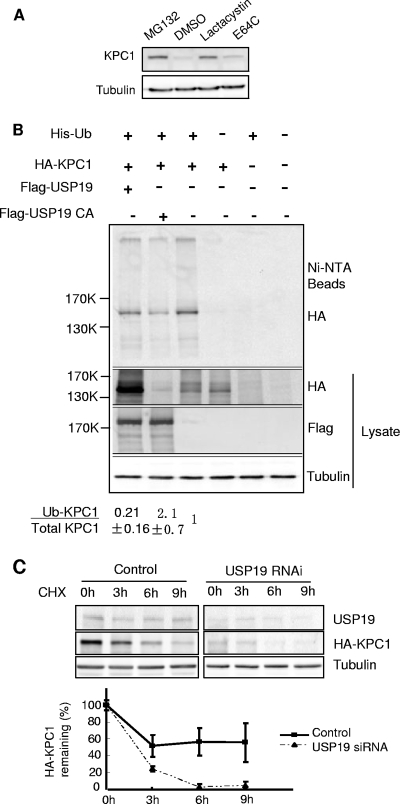
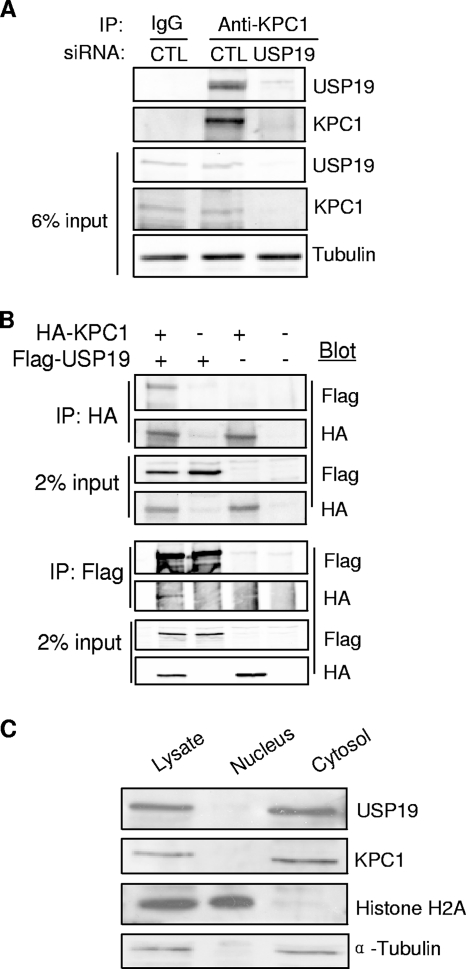
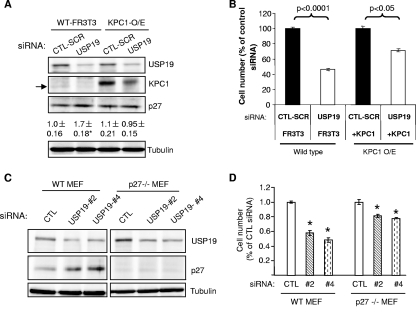
p27(Kip1) is a cyclin-dependent kinase inhibitor that regulates the G(1)/S transition. Increased degradation of p27(Kip1) is associated with cellular transformation. Previous work demonstrated that the ubiquitin ligases KPC1/KPC2 and SCF(Skp2) ubiquitinate p27(Kip1) in G(1) and early S, respectively. The regulation of these ligases remains unclear. We report here that the USP19 deubiquitinating enzyme interacts with and stabilizes KPC1, thereby modulating p27(Kip1) levels and cell proliferation. Cells depleted of USP19 by RNA interference exhibited an inhibition of cell proliferation, progressing more slowly from G(0)/G1 to S phase, and accumulated p27(Kip1). This increase in p27(Kip1) was associated with normal levels of Skp2 but reduced levels of KPC1. The overexpression of KPC1 or the use of p27(-/-) cells inhibited significantly the growth defect observed upon USP19 depletion. KPC1 was ubiquitinated in vivo and stabilized by proteasome inhibitors and by overexpression of USP19, and it also coimmunoprecipitated with USP19. Our results identify USP19 as the first deubiquitinating enzyme that regulates the stability of a cyclin-dependent kinase inhibitor and demonstrate that progression through G(1) to S phase is, like the metaphase-anaphase transition, controlled in a hierarchical, multilayered fashion.
Figures
References
-
- Bloom, J., and M. Pagano. 2003. Deregulated degradation of the cdk inhibitor p27 and malignant transformation. Semin. Cancer Biol. 1341-47. - PubMed
-
- Burrows, J. F., M. J. McGrattan, A. Rascle, M. Humbert, K. H. Baek, and J. A. Johnston. 2004. DUB-3, a cytokine-inducible deubiquitinating enzyme that blocks proliferation. J. Biol. Chem. 27913993-14000. - PubMed
-
- Carrano, A. C., E. Eytan, A. Hershko, and M. Pagano. 1999. SKP2 is required for ubiquitin-mediated degradation of the CDK inhibitor p27. Nat. Cell Biol. 1193-199. - PubMed
-
- Crosas, B., J. Hanna, D. S. Kirkpatrick, D. P. Zhang, Y. Tone, N. A. Hathaway, C. Buecker, D. S. Leggett, M. Schmidt, R. W. King, S. P. Gygi, and D. Finley. 2006. Ubiquitin chains are remodeled at the proteasome by opposing ubiquitin ligase and deubiquitinating activities. Cell 1271401-1413. - PubMed
-
- Deng, C., P. Zhang, J. W. Harper, S. J. Elledge, and P. Leder. 1995. Mice lacking p21CIP1/WAF1 undergo normal development, but are defective in G1 checkpoint control. Cell 82675-684. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous