

Formation and longevity of chimeric and duplicate genes in Drosophila melanogaster
- PMID: 19015547
- PMCID: PMC2621179
- DOI: 10.1534/genetics.108.091538
Formation and longevity of chimeric and duplicate genes in Drosophila melanogaster
Abstract
Historically, duplicate genes have been regarded as a major source of novel genetic material. However, recent work suggests that chimeric genes formed through the fusion of pieces of different genes may also contribute to the evolution of novel functions. To compare the contribution of chimeric and duplicate genes to genome evolution, we measured their prevalence and persistence within Drosophila melanogaster. We find that approximately 80.4 duplicates form per million years, but most are rapidly eliminated from the genome, leaving only 4.1% to be preserved by natural selection. Chimeras form at a comparatively modest rate of approximately 11.4 per million years but follow a similar pattern of decay, with ultimately only 1.4% of chimeras preserved. We propose two mechanisms of chimeric gene formation, which rely entirely on local, DNA-based mutations to explain the structure and placement of the youngest chimeric genes observed. One involves imprecise excision of an unpaired duplication during large-loop mismatch repair, while the other invokes a process akin to replication slippage to form a chimeric gene in a single event. Our results paint a dynamic picture of both chimeras and duplicate genes within the genome and suggest that chimeric genes contribute substantially to genomic novelty.
Figures
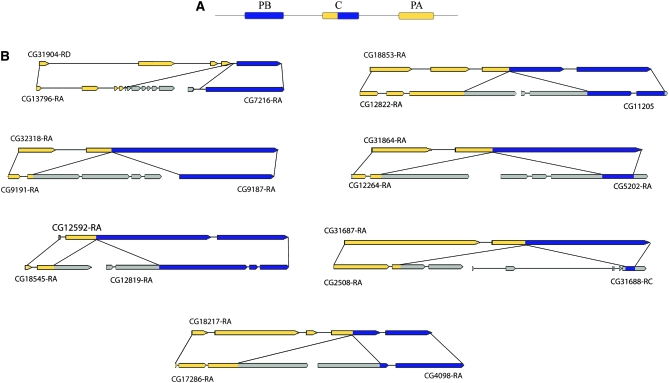
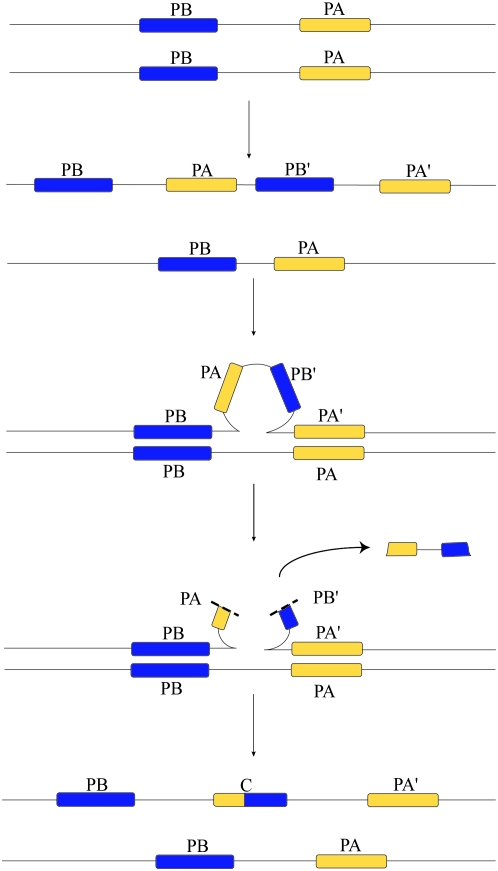
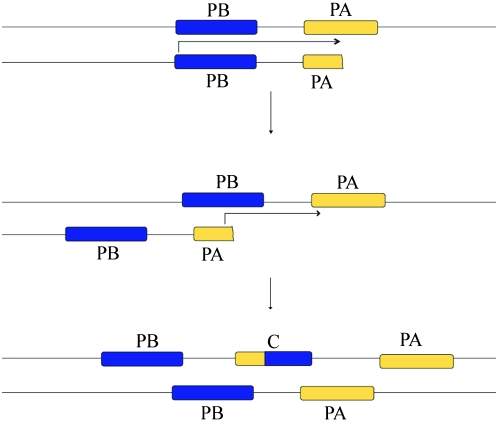
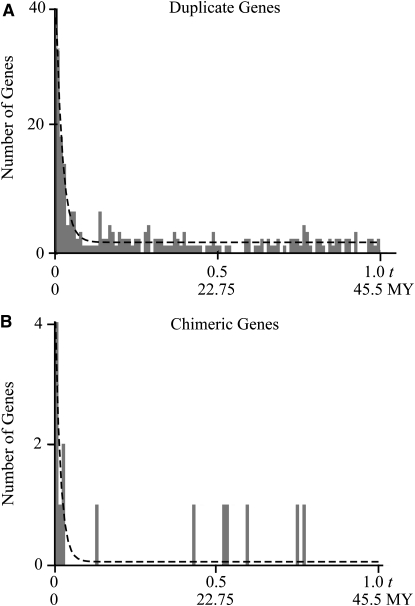
References
-
- Adams, M. D., S. E. Celniker, R. A. Holt, C. A. Evans, J. D. Gocayne et al., 2000. The genome sequence of Drosophila melanogaster. Science 287 2185–2195. - PubMed
-
- Altschul, S. F., W. Gish, W. Miller, E. W. Myers and D. J. Lipman, 1990. Basic local alignment search tool. J. Mol. Biol. 215 403–410. - PubMed
-
- Alvarez, A. D., W. Shi, B. A. Wilson and J. B. Skeath, 2003. Pannier and pointedP2 act sequentially to regulate Drosophila heart development. Development 130 3015–3026. - PubMed
-
- Brown, S., and J. Castelli-Gair Hombria, 2000. Drosophila grain encodes a GATA transcription factor required for cell rearrangement during morphogenesis. Development 127 4867–4876. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
