

The Caenorhabditis elegans ing-3 gene regulates ionizing radiation-induced germ-cell apoptosis in a p53-associated pathway
- PMID: 19015549
- PMCID: PMC2644941
- DOI: 10.1534/genetics.107.080515
The Caenorhabditis elegans ing-3 gene regulates ionizing radiation-induced germ-cell apoptosis in a p53-associated pathway
Abstract
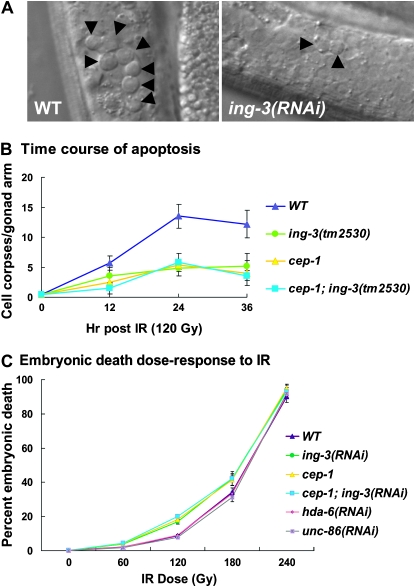
The inhibitor of growth (ING) family of type II tumor suppressors are encoded by five genes in mammals and by three genes in Caenorhabditis elegans. All ING proteins contain a highly conserved plant homeodomain (PHD) zinc finger. ING proteins are activated by stresses, including ionizing radiation, leading to the activation of p53. ING proteins in mammals and yeast have recently been shown to read the histone code in a methylation-sensitive manner to regulate gene expression. Here we identify and characterize ing-3, the C. elegans gene with the highest sequence identity to the human ING3 gene. ING-3 colocalizes with chromatin in embryos, the germline, and somatic cells. The ing-3 gene is part of an operon but is also transcribed from its own promoter. Both ing-3(RNAi) and ing-3 mutant strains demonstrate that the gene likely functions in concert with the C. elegans p53 homolog, cep-1, to induce germ-cell apoptosis in response to ionizing radiation. Somatically, the ing-3 mutant has a weak kinker uncoordinated (kinker Unc) phenotype, indicating a possible neuronal function.
Figures



References
-
- Bienz, M., 2006. The PHD finger, a nuclear protein-interaction domain. Trends Biochem. Sci. 31 35–40. - PubMed
-
- Blumenthal, T., and K. Steward, 1997. RNA processing and gene structure, pp. 117–146 in C. elegans II, edited by D. L. Riddle, T. Blumenthal, B. J. Meyer and J. R. Priess. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
