

Identification of mitochondrial F(1)F(0)-ATP synthase interacting with galectin-3 in colon cancer cells
- PMID: 19016746
- PMCID: PMC11160105
- DOI: 10.1111/j.1349-7006.2008.00901.x
Identification of mitochondrial F(1)F(0)-ATP synthase interacting with galectin-3 in colon cancer cells
Abstract
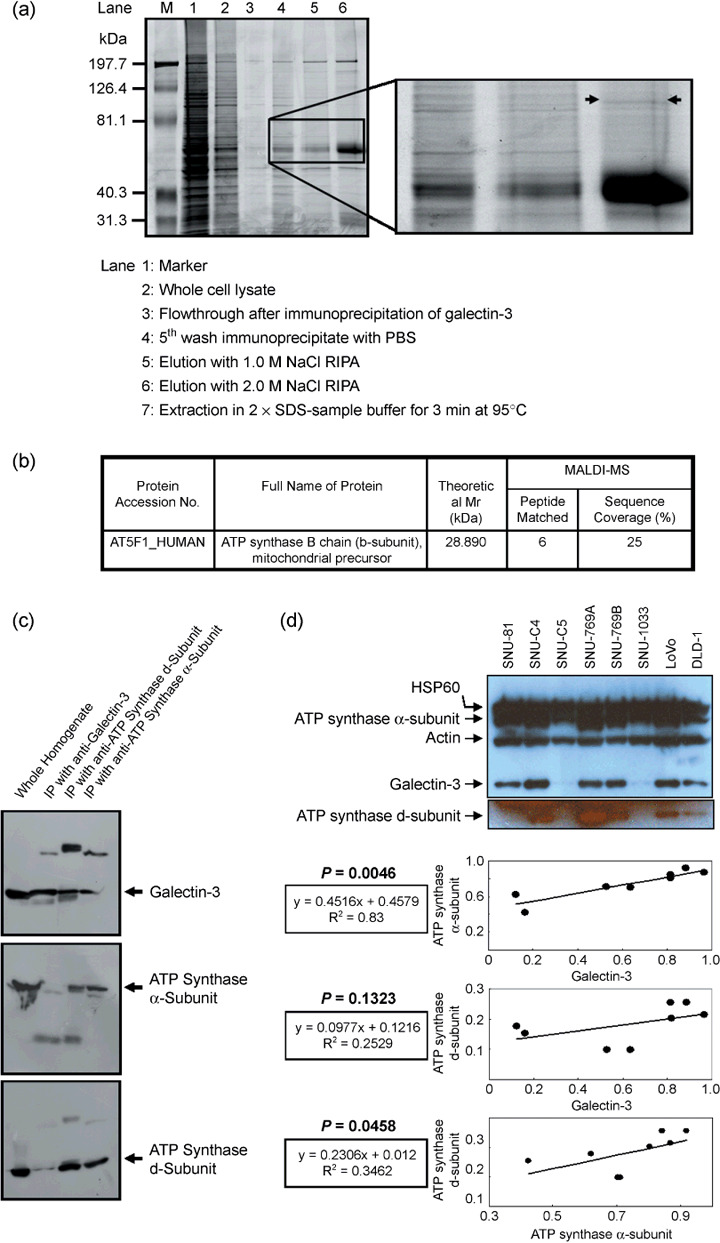
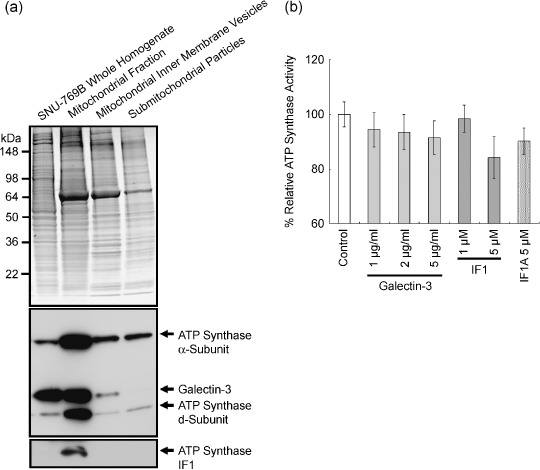
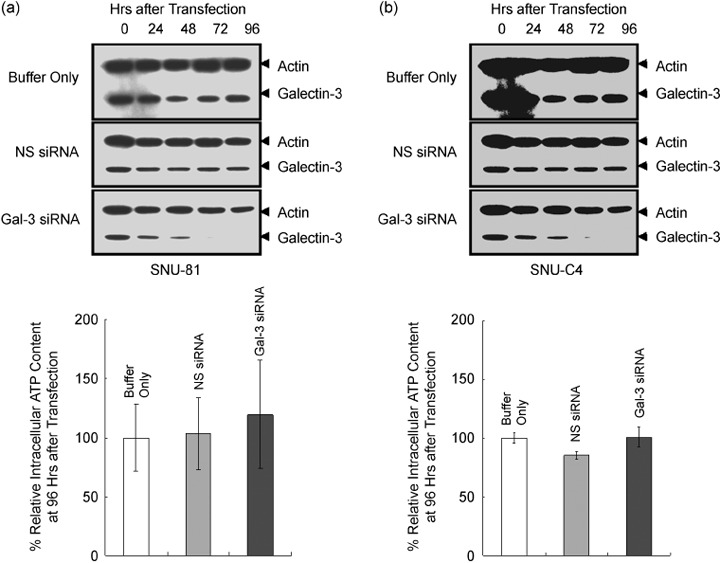
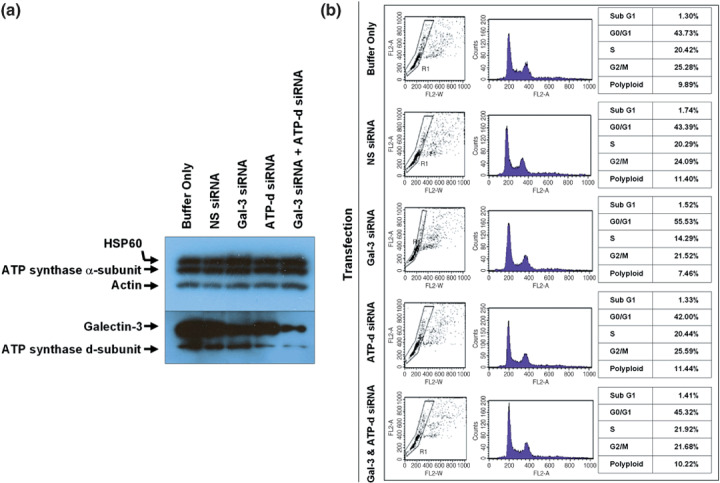
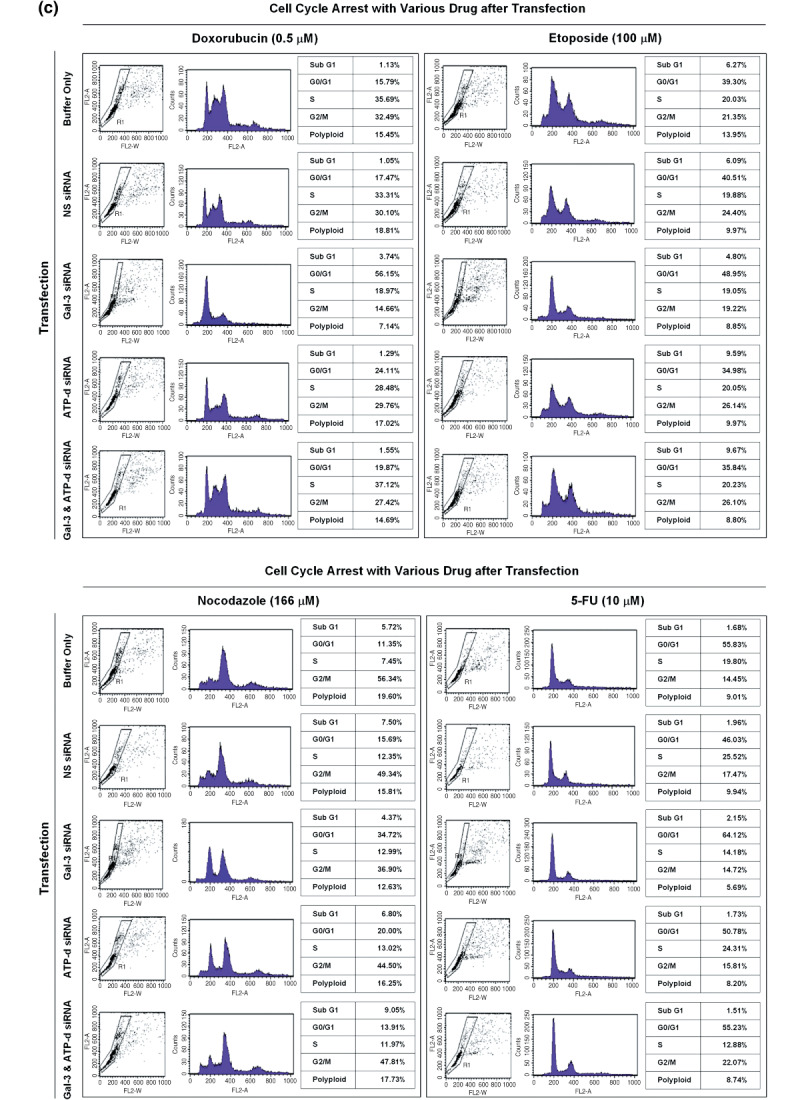
To evaluate the effect of galectin-3 in cell cycle regulation of colon cancer cells, we looked for binding molecules interacting with galectin-3 and examined the changes in cell cycle by suppressing galectin-3 and the binding molecule. To identify target molecules interacting with galectin-3, we analyzed immunoprecipitate of the anti-galectin-3 antibody obtained from human colon cancer cell line, using matrix-assisted laser desorption ionization-mass spectrometry. We validated subcellular localization of galectin-3 and ATP synthase identified, and ATP synthase activity was determined in the presence of galectin-3. Cell cycle regulation was monitored after galectin-3 siRNA transfection. ATP synthase b-subunit was identified in immunoprecipitate of the anti-galectin-3 antibody. Galectin-3 and ATP synthase were co-isolated in the inner membrane vesicles of mitochondria. Galectin-3 has an inhibitory activity against ATP synthase, and intracellular ATP content showed increasing tendency after galectin-3 suppression. Suppression of galectin-3 resulted in G0/G1 progression of human colon cancer cells arrested at S, S/G2 and G2/M phase in the presence of doxorubicin, and etoposide or nocodazole, respectively. Compared to cells in which ATP synthase d-subunit was suppressed alone, sub-G1 fraction caused by etoposide or nocodazole was decreased in cells with galectin-3 suppression alone. In conclusion, galectin-3 co-localized with ATP synthase in the inner membrane of mitochondria and has an inhibitory effect on ATP synthase in human colon cancer cells. In the presence of cell cycle synchronizing drugs, doxorubicin, etoposide, or nocodazole, suppression of galectin-3 induced cell cycle progression to G0/G1 phase.
Figures
References
-
- Barondes SH, Castronovo V, Cooper DN et al . Galectins: a family of animal β‐galactoside‐binding lectins. Cell 1994; 76: 597–8. - PubMed
-
- Dumic J, Dabelic S, Flogel M. Galectin‐3: an open‐ended story. Biochimica et Biophysica Acta 2006; 1760: 616–35. - PubMed
-
- Ruebel KH, Jin L, Qian X et al . Effects of DNA methylation on galectin‐3 expression in pituitary tumors. Cancer Res 2005; 65: 1136–40. - PubMed
-
- Paz A, Haklai R, Elad‐Sfadia G et al . Galectin‐1 binds oncogenic H‐Ras to mediate Ras membrane anchorage and cell transformation. Oncogene 2001; 20: 7486–93. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
