

A novel IFN regulatory factor 3-dependent pathway activated by trypanosomes triggers IFN-beta in macrophages and fibroblasts
- PMID: 19017982
- PMCID: PMC2597000
- DOI: 10.4049/jimmunol.181.11.7917
A novel IFN regulatory factor 3-dependent pathway activated by trypanosomes triggers IFN-beta in macrophages and fibroblasts
Abstract
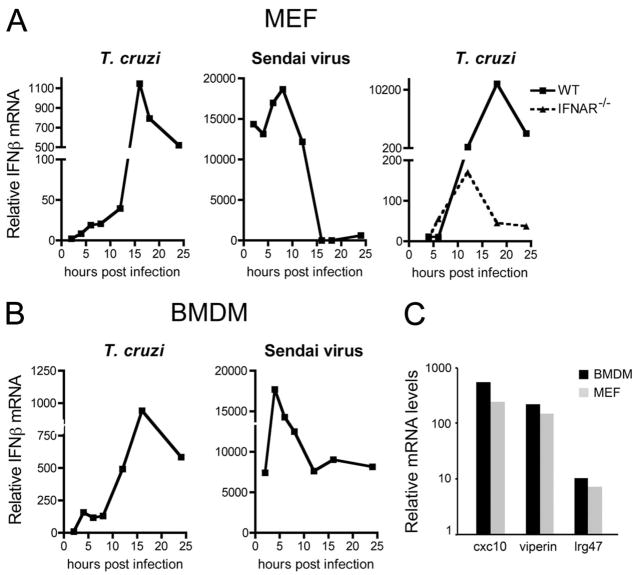
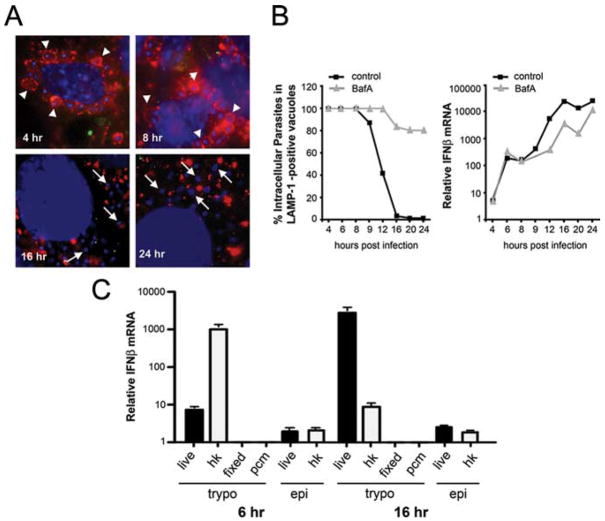
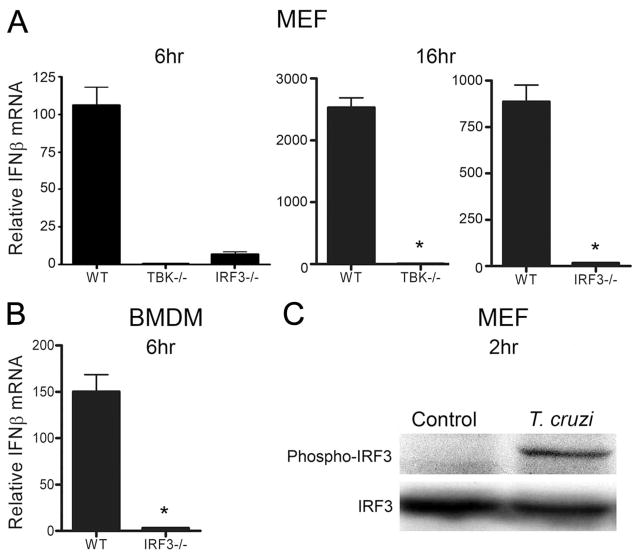
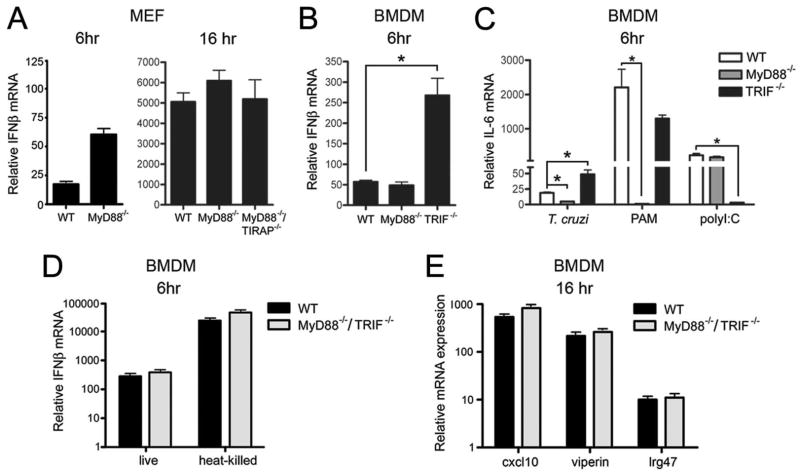
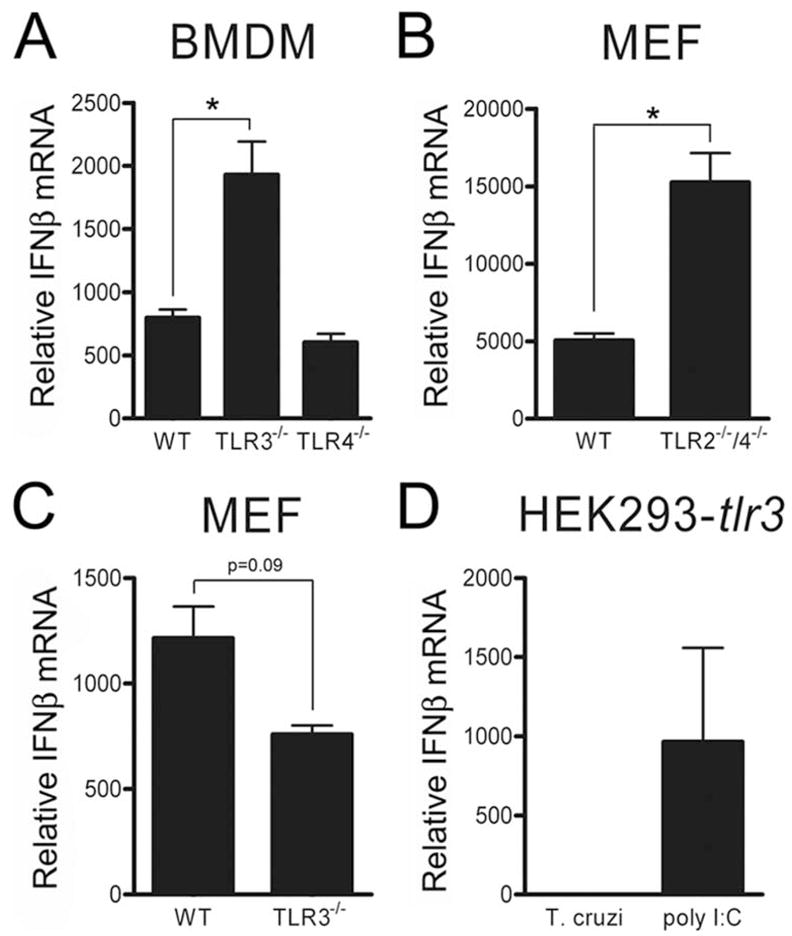
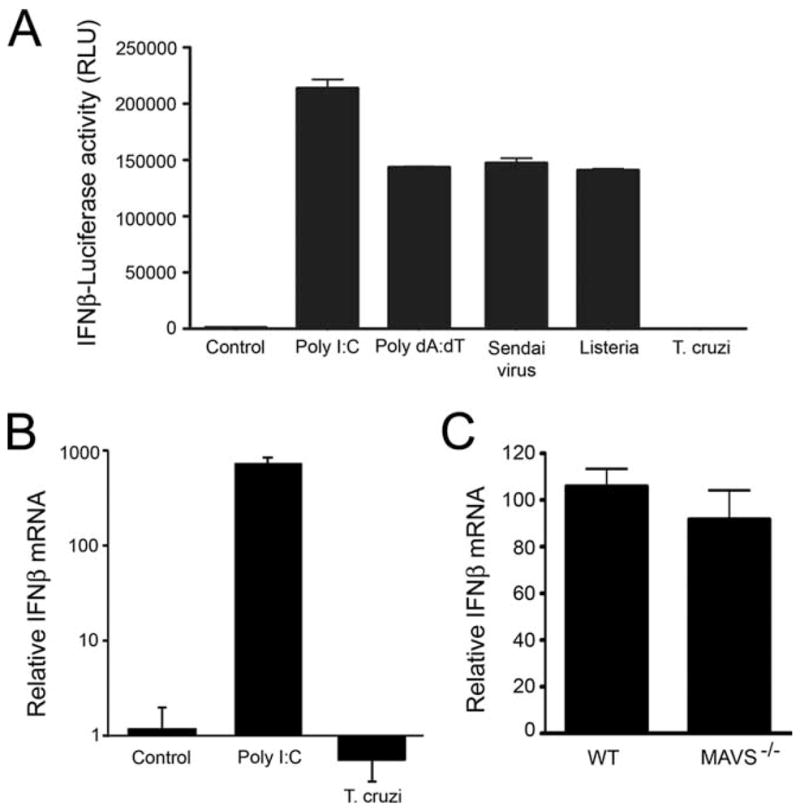
Innate immune recognition of intracellular pathogens involves both extracellular and cytosolic surveillance mechanisms. The intracellular protozoan parasite Trypanosoma cruzi triggers a robust type I IFN response in both immune and nonimmune cell types. In this study, we report that signaling through TBK1 and IFN regulatory factor 3 is required for T. cruzi-mediated expression of IFN-beta. The TLR adaptors MyD88 and TRIF, as well as TLR4 and TLR3, were found to be dispensable, demonstrating that T. cruzi induces IFN-beta expression in a TLR-independent manner. The potential role for cytosolic dsRNA sensing pathways acting through RIG-I and MDA5 was ruled out because T. cruzi was shown to trigger robust expression of IFN-beta in macrophages lacking the MAVS/IPS1/VISA/CARDif adaptor protein. The failure of T. cruzi to activate HEK293-IFN-beta-luciferase cells, which are highly sensitive to cytosolic triggers of IFN-beta expression including Listeria, Sendai virus, and transfected dsRNA and dsDNA, further indicates that the parasite does not engage currently recognized cytosolic surveillance pathways. Together, these findings identify the existence of a novel TLR-independent pathogen-sensing mechanism in immune and nonimmune cells that converges on TBK1 and IFN regulatory factor 3 for activation of IFN-beta gene expression.
Conflict of interest statement
Disclosures
The authors have no financial conflict of interest.
Figures
References
-
- Bogdan C, Mattner J, Schleicher U. The role of type I interferons in non-viral infections. Immunol Rev. 2004;202:33–48. - PubMed
-
- Noppert SJ, Fitzgerald KA, Hertzog PJ. The role of type I interferons in TLR responses. Immunol Cell Biol. 2007;85:446–457. - PubMed
-
- Akira S, Uematsu S, Takeuchi O. Pathogen recognition and innate immunity. Cell. 2006;124:783–801. - PubMed
-
- Yoneyama M, Kikuchi M, Natsukawa T, Shinobu N, Imaizumi T, Miyagishi M, Taira K, Akira S, Fujita T. The RNA helicase RIG-I has an essential function in double-stranded RNA-induced innate antiviral responses. Nat Immunol. 2004;5:730–737. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
