

Multidrug resistance-associated transporter 2 regulates mucosal inflammation by facilitating the synthesis of hepoxilin A3
- PMID: 19017997
- PMCID: PMC2596662
- DOI: 10.4049/jimmunol.181.11.8044
Multidrug resistance-associated transporter 2 regulates mucosal inflammation by facilitating the synthesis of hepoxilin A3
Abstract
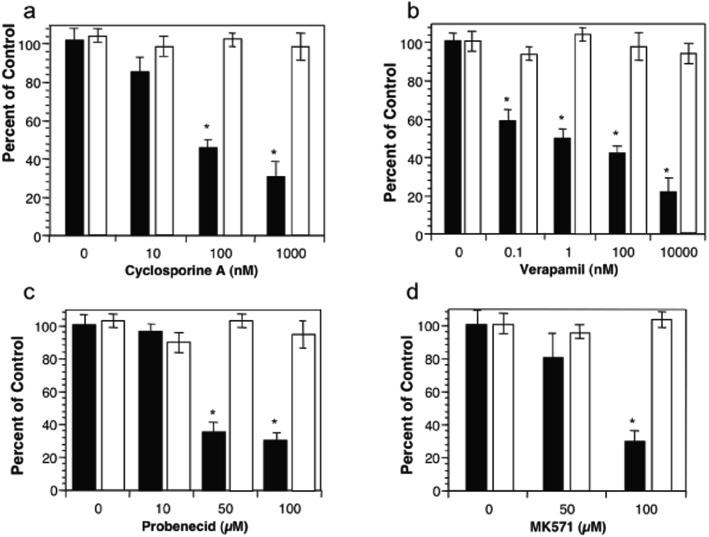
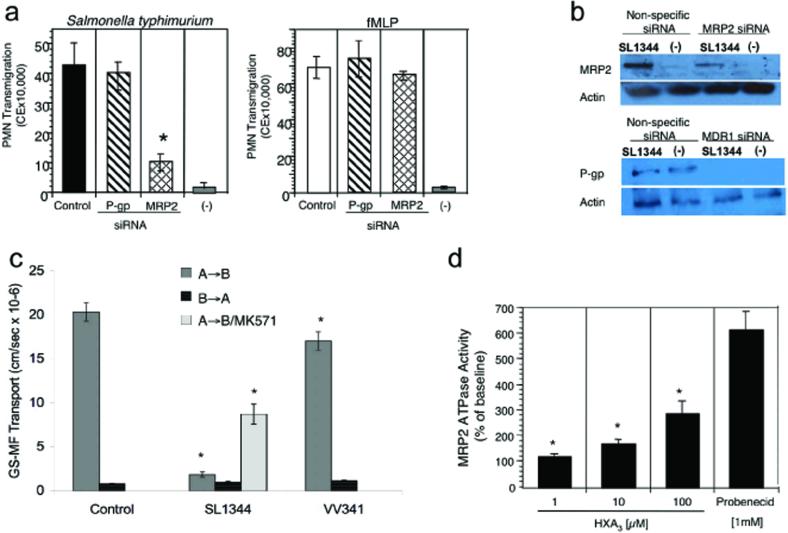
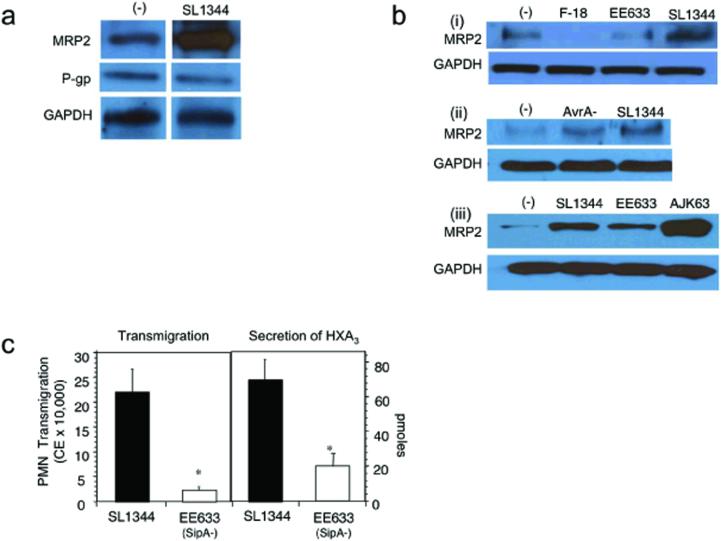
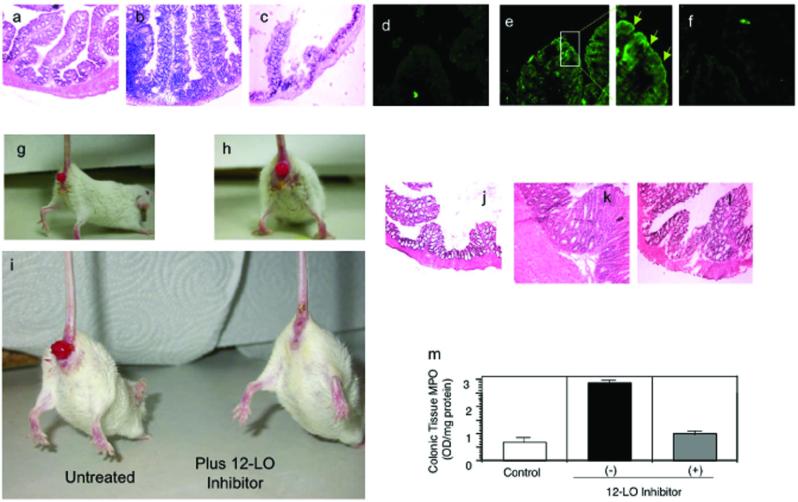
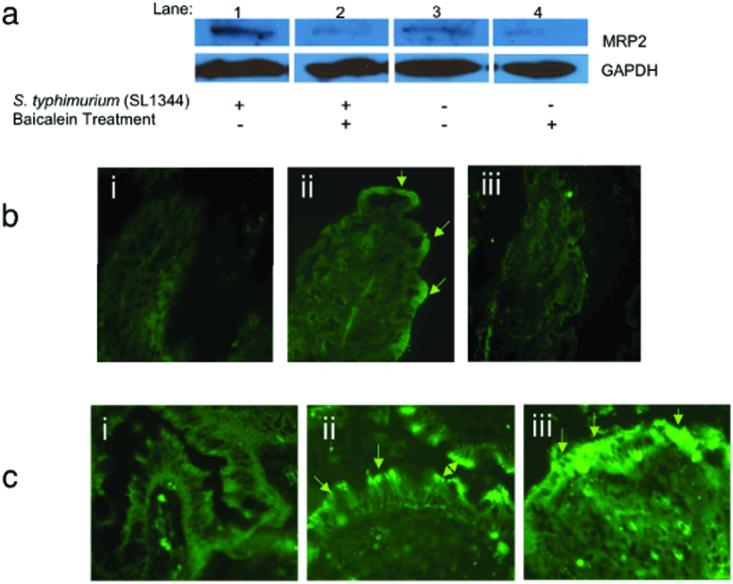
Neutrophil transmigration across mucosal surfaces contributes to dysfunction of epithelial barrier properties, a characteristic underlying many mucosal inflammatory diseases. Thus, insight into the directional movement of neutrophils across epithelial barriers will provide important information relating to the mechanisms of such inflammatory disorders. The eicosanoid hepoxilin A(3), an endogenous product of 12-lipoxygenase activity, is secreted from the apical surface of the epithelial barrier and establishes a chemotactic gradient to guide neutrophils from the submucosa across epithelia to the luminal site of an inflammatory stimulus, the final step in neutrophil recruitment. Currently, little is known regarding how hepoxilin A(3) is secreted from the intestinal epithelium during an inflammatory insult. In this study, we reveal that hepoxilin A(3) is a substrate for the apical efflux ATP-binding protein transporter multidrug resistance-associated protein 2 (MRP2). Moreover, using multiple in vitro and in vivo models, we show that induction of intestinal inflammation profoundly up-regulates apical expression of MRP2, and that interfering with hepoxilin A(3) synthesis and/or inhibition of MRP2 function results in a marked reduction in inflammation and severity of disease. Lastly, examination of inflamed intestinal epithelia in human biopsies revealed up-regulation of MRP2. Thus, blocking hepoxilin A(3) synthesis and/or inhibiting MRP2 may lead to the development of new therapeutic strategies for the treatment of epithelial-associated inflammatory conditions.
Figures
Similar articles
-
Hepoxilin A(3) facilitates neutrophilic breach of lipoxygenase-expressing airway epithelial barriers.J Immunol. 2012 Nov 15;189(10):4960-9. doi: 10.4049/jimmunol.1201922. Epub 2012 Oct 8. J Immunol. 2012. PMID: 23045615 Free PMC article. Clinical Trial.
-
Transporters MRP1 and MRP2 Regulate Opposing Inflammatory Signals To Control Transepithelial Neutrophil Migration during Streptococcus pneumoniae Lung Infection.mSphere. 2018 Jul 5;3(4):e00303-18. doi: 10.1128/mSphere.00303-18. mSphere. 2018. PMID: 29976647 Free PMC article.
-
Identification of hepoxilin A3 in inflammatory events: a required role in neutrophil migration across intestinal epithelia.Proc Natl Acad Sci U S A. 2004 May 11;101(19):7421-6. doi: 10.1073/pnas.0400832101. Epub 2004 May 3. Proc Natl Acad Sci U S A. 2004. PMID: 15123795 Free PMC article.
-
Bacterial-induced hepoxilin A3 secretion as a pro-inflammatory mediator.FEBS J. 2007 Jul;274(14):3513-3518. doi: 10.1111/j.1742-4658.2007.05911.x. Epub 2007 Jul 2. FEBS J. 2007. PMID: 17608718 Review.
-
The role of neutrophils during intestinal inflammation.Mucosal Immunol. 2012 Jul;5(4):354-66. doi: 10.1038/mi.2012.24. Epub 2012 Apr 11. Mucosal Immunol. 2012. PMID: 22491176 Review.
Cited by
-
The fimbriae of enteroaggregative Escherichia coli induce epithelial inflammation in vitro and in a human intestinal xenograft model.J Infect Dis. 2012 Sep 1;206(5):714-22. doi: 10.1093/infdis/jis417. Epub 2012 Jun 21. J Infect Dis. 2012. PMID: 22723643 Free PMC article.
-
Mucosal Inflammatory Response to Salmonella typhimurium Infection.Front Immunol. 2014 Jul 4;5:311. doi: 10.3389/fimmu.2014.00311. eCollection 2014. Front Immunol. 2014. PMID: 25071772 Free PMC article. Review.
-
Intermediate role of gut microbiota in vitamin B nutrition and its influences on human health.Front Nutr. 2022 Dec 13;9:1031502. doi: 10.3389/fnut.2022.1031502. eCollection 2022. Front Nutr. 2022. PMID: 36583209 Free PMC article. Review.
-
Altered Expression of Small Intestinal Drug Transporters and Hepatic Metabolic Enzymes in a Mouse Model of Familial Alzheimer's Disease.Mol Pharm. 2018 Sep 4;15(9):4073-4083. doi: 10.1021/acs.molpharmaceut.8b00500. Epub 2018 Aug 22. Mol Pharm. 2018. PMID: 30074800 Free PMC article.
-
Illuminating dynamic neutrophil trans-epithelial migration with micro-optical coherence tomography.Sci Rep. 2017 Apr 3;8:45789. doi: 10.1038/srep45789. Sci Rep. 2017. PMID: 28368012 Free PMC article.
References
-
- Pace-Asciak CR, Reynaud D, Demin P, Nigam S. The hepoxilins. A review. Adv Exp Med Biol. 1999;447:123–132. - PubMed
-
- Nigam S, Zafiriou MP, Deva R, Ciccoli R, Roux-Van der Merwe R. Structure, biochemistry and biology of hepoxilins: an update. Febs J. 2007;274:3503–3512. - PubMed
-
- Nigam S, Patabhiraman S, Ciccoli R, Ishdorj G, Schwarz K, Petrucev B, Kuhn H, Haeggstrom JZ. The rat leukocyte-type 12-lipoxygenase exhibits an intrinsic hepoxilin A3 synthase activity. J Biol Chem. 2004;279:29023–29030. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
